




El uracilo en el ADN: ¿error o señal? Understand article
Traducido por Maialen Ruiz Prada. El uracilo es conocido como una de las bases que forman parte del RNA pero ¿por qué no se utiliza en el DNA?¿o sí se utiliza? Angéla Békési y Beáta G Vértessy investigaan.
Timina frente a uracilo
Nuestra información genética se almacena en forma de DNA, utilizando un alfabeto de cuatro letras. Las cuatro “letras” corresponden a las cuatro bases químicas que cada bloque de construcción de DNA (llamado nucleótido) puede tener: adenina (A), timina (T), citosina (C) y guanina (G, ver Figura 1). Como mostró el por todos conocido descubrimiento de James Watson y Francis Crick, el DNA forma una doble hélice en la que las cuatro bases siempre se emparejan de la misma manera, a través de enlaces de hidrógeno específicos: la adenina se une a la timina y la guanina a la citosina (ver Figuras 2 y 3).
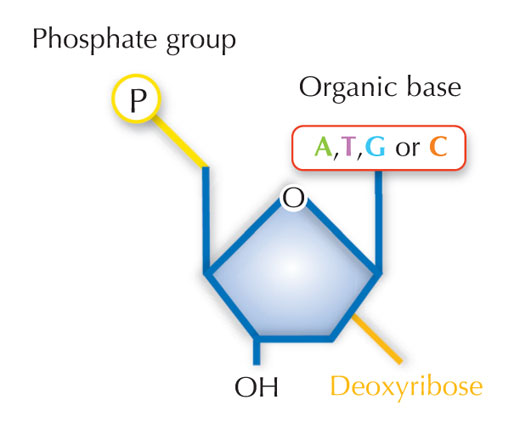
Imagen cortesía de Nicola Graf
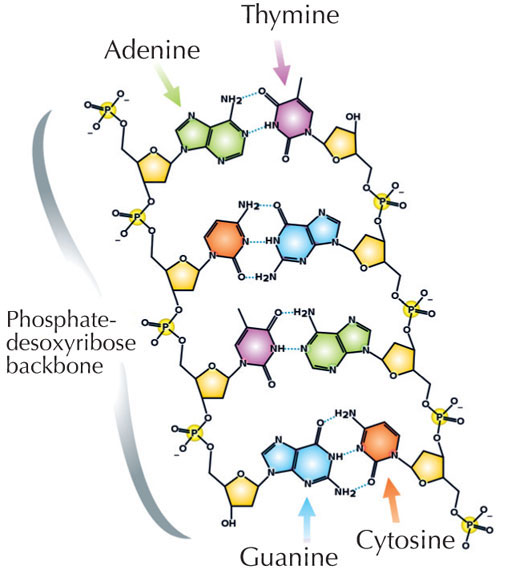
Imagen cortesía de Madeleine Price Ball. Fuente de la imagen: Wikimedia Commons
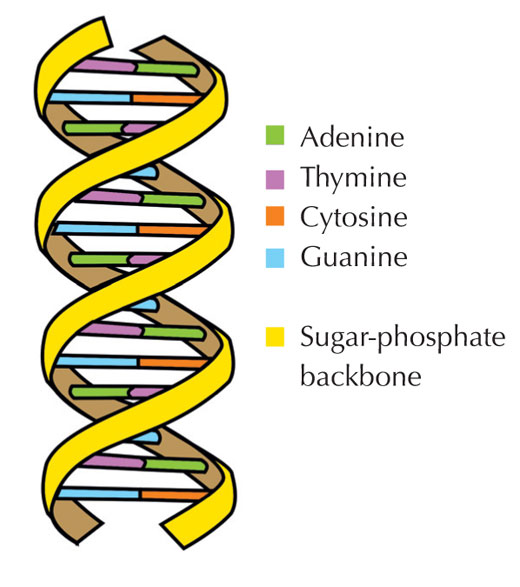
Imagen cortesía de Forluvoft. Fuente de la imagen: Wikimedia Commons
Sin embargo, hay una quinta letra alternativa: el uracilo (U), que forma el mismo tipo de enlaces de hidrógeno con la adenina (ver Figura 4). Pero a pesar de que el uracilo es muy común en el RNA, no es así en el caso del DNA, donde en vez de uracilo se utiliza timina. ¿Por qué ocurre esto?
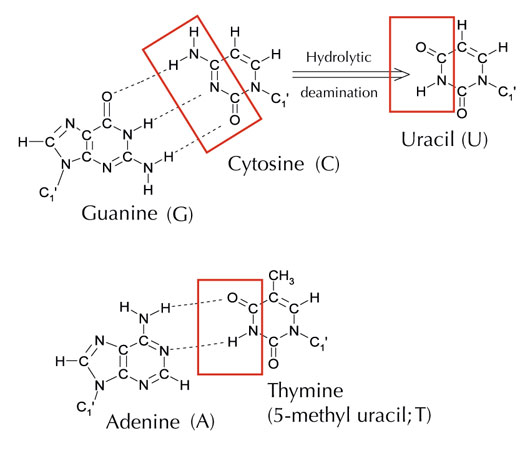
Imagen cortesía de Angéla Békési
Químicamente, la timina es una molécula de uracilo con un grupo metilo adicional. ¿Cuál sería la ventaja, en términos evolutivos, de la utilización de este bloque de construcción más complejo en el DNA? La respuesta puede estar en cómo corrigen las células los daños en el DNA.
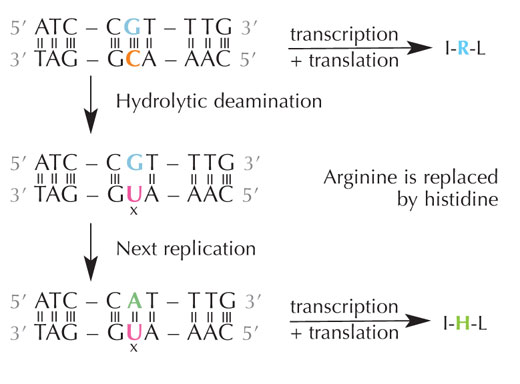
hidrolítica de la citosina
puede cambiar los
aminoácidos codificados por
la secuencia . Haga clic para
ampliar la imagen
Imagen cortesía de Nicola Graf
La citosina se puede convertir espontáneamente en uracilo a través de un proceso llamado desaminación hidrolítica (ver Figura 4). Cuando esto ocurre, la guanina que estaba unida inicialmente a esa molécula de citosina pasa a estar enfrente del uracilo (recordemos que el uracilo normalmente se une a la adenina). La próxima vez que la célula replique su DNA, la posición opuesta a esta molécula de uracilo se llenará con una adenina en lugar de con la guanina que debería estar ahí, alterando el mensaje que codifica esta sección de DNA (ver Figura 5). Este proceso de desaminación de la citosina es uno de los tipos más comunes de daños en el DNA, pero normalmente se corrige de manera eficaz. ¿Cómo hace esto la célula?
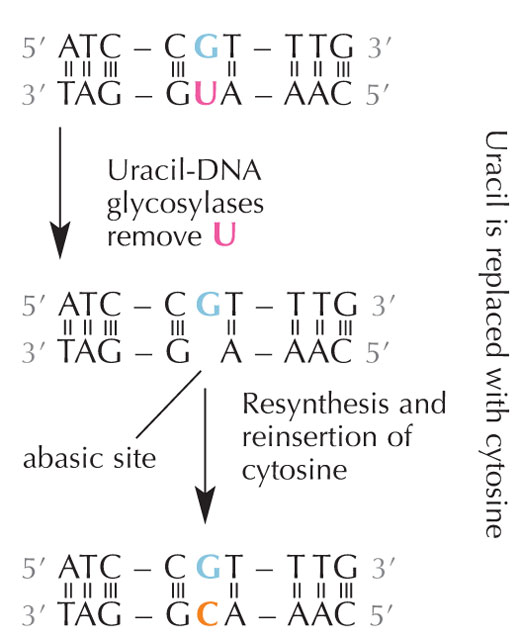
desaminación hidrolítica.
Haga clic para ampliar la
imagen
Imagen cortesía de Nicola Graf
Las células tienen un sistema de reparación que puede detectar cuándo hay un uracilo donde debería haber una citosina y corregir el error antes de que se replique y se transmita. La compleja maquinaria encargada de hacer esto consiste en un conjunto de varias enzimas: primero, las uracil-DNA glicosilasas reconocen el uracilo y lo quitan del DNA. Después, varias enzimas contribuyen a la eliminación y posterior síntesis de la parte dañada del DNA, durante el cual el sitio abásico (“vacío”) en el DNA se rellena con una citosina (ver Figura 6).
Sin embargo, la forma más común de la uracil-DNA glicosilasa no puede distinguir con qué base está emparejada el uracilo, es decir, si el uracilo tenía que estar allí (emparejado con adenina) o si se trata de una citosina mutada (con su pareja, una guanina). La enzima cortaría los dos tipos de uracilo y claramente esto causaría problemas. Se cree que la solución a este problema potencial ha consistido en la evolución de un mecanismo por el cual los uracilos “correctos” (emparejados con adenina) eran marcados con un grupo metilo, dando lugar a la timina. De esta manera, si la maquinaria de la célula encontraba un uracilo lo quitaba y reparaba el daño, pero si se encontraba un uracilo marcado con un metilo, es decir, una timina (ver Figura 4), lo dejaba. Con el tiempo, por lo tanto, la timina desplazó al uracilo en el DNA, y ahora la mayoría de las células solo utilizan uracilo en el RNA.
¿Por qué se mantuvo el uracilo en el RNA? El RNA es más efímero que el DNA y, con pocas excepciones, no es un repositorio de la información genética a largo plazo, por lo que las moléculas de citosina que se convierten espontáneamente en uracilo en el RNA no son una gran amenaza para la célula. Por lo tanto, probablemente no existió demasiada presión evolutiva para reemplazar el uracilo por la más compleja (y probablemente más costosa) timina en el RNA.
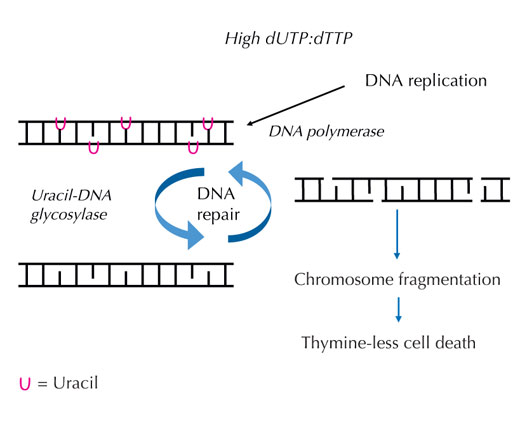
proporción dUTP:dTTP, la
DNA polimerasa incorporará
con frecuencia uracilo en
lugar de timina durante la
replicación y la reparación. La
uracil-DNA glicosilasa quita
el uracilo e inicia la
reparación implicando cortes
en la cadena del DNA como
paso intermedio. Sin
embargo, esta síntesis
reparadora puede
reintroducir moléculas de
uracilo, provocando que ese
ciclo de reparación sea inútil.
Con el tiempo, el sistema se
sobrecarga y se produce la
fragmentación cromosómica,
lo que lleva a la muerte
celular. Haga clic para
ampliar la imagen
Imagen cortesía de Angéla
Békési
Muerte celular por falta de timina
Durante la síntesis del DNA, las enzimas DNA polimerasa (que catalizan la síntesis) no son capaces de distinguir entre timina y uracilo. Solo comprueban si los enlaces de hidrógeno se han formado correctamente, es decir, si los pares de bases se han emparejado bien. A estas enzimas no les importa si es la timina o el uracilo el que se une a la adenina. Normalmente, la cantidad de desoxiuridina trifosfato (dUTP, una fuente de uracilo) en la célula se mantiene muy baja en comparación con los niveles de desoxitimidina trifosfato (dTTP, una fuente de timina), impidiendo la incorporación de uracilo durante la síntesis del DNA.
Si algo perturba esta estricta regulación y la proporción de dUTP frente a dTTP se eleva, la cantidad de uracilo que se incorpora al DNA de forma incorrecta también aumenta. El sistema de reparación que, a diferencia de las DNA polimerasas, puede distinguir uracilo de timina, intenta a continuación quitar el uracilo con la ayuda de la uracil-DNA glicosilasa y volver a sintetizar el DNA, lo que implica cortar temporalmente el esqueleto del DNA. Sin embargo, si la proporción de dUTP frente a dTTP sigue siendo elevada, esta segunda síntesis puede volver a incorporar uracilo en lugar de timina. Al final, si estos cortes temporales en el DNA se producen uno tras otro y muy seguidos, este ciclo conducirá a roturas de la cadena del DNA y a la fragmentación cromosómica (ver Figura 7). Esto da lugar a un tipo específico de muerte celular programada, denominada muerte celular por falta de timina.
El proceso de la muerte celular por falta de timina puede ser aprovechado deliberadamente en el tratamiento del cáncer. Debido a que las células cancerosas proliferan a una velocidad muy alta en comparación con las células normales, sintetizan una mayor cantidad de DNA en un periodo de tiempo dado y, por lo tanto, requieren grandes cantidades de dUTP. Al aumentar la proporción de dUTP frente a dTTP, estas células cancerosas pueden ser seleccionadas y eliminadas de forma selectiva.
El uracilo todavía existe en el DNA
Aunque la mayoría de las células utilizan uracilo para el RNA y timina para el DNA, existen excepciones. Algunos organismos tienen uracilo en lugar de timina en todo su DNA, y otros organismos tienen uracilo en solo una parte de su DNA. ¿Cuál podría ser la ventaja evolutiva de eso? Echemos un vistazo a algunos ejemplos.
El uracilo en el DNA viral
un fago infectando una célula
bacteriana
Imagen cortesía de cdascher /
iStockphoto
Se conocen dos especies de fagos (virus que infectan bacterias) que tienen genomas de DNA con uracilo y sin timina. Todavía no sabemos si estos fagos son representantes de una antigua forma de vida que nunca evolucionó a tener timina en el DNA o si sus genomas con uracilo son una estrategia de evolución reciente. Tampoco sabemos por qué utilizan estos fagos uracilo en lugar de timina, pero esto puede desempeñar un papel esencial en el ciclo de vida de estos virus. Si ese es el caso, tendría sentido que los virus se aseguraran de que el uracilo no se sustituye por timina en su DNA. Y, de hecho, se ha demostrado que uno de estos fagos tiene un gen que codifica para una proteína específica que inhibe la uracil-DNA glicosilasa del huésped, evitando así que las enzimas del huésped reparen el uracilo del genoma viral.
La muerte celular programada en el ciclo de vida de los insectos

hormigas carecen de la
enzima capaz de eliminar el
uracilo de su DNA
Imagen cortesía de spxChrome
/ iStockphoto y Nicola Graf
El DNA con uracilo también parece desempeñar un papel en el desarrollo de los endopterigotos, que son insectos que pasan por una fase de pupa durante su ciclo de vida (por ejemplo, las hormigas y los mariposas lo hacen pero los saltamontes y las termitas no). Estos insectos carecen del gen principal de la uracil-DNA glicosilasa, que de ser producida eliminaría el uracilo de su DNA.
Por otra parte, nuestras propias investigaciones han demostrado que en las larvas de la mosca de la fruta Drosophila melanogaster la proporción de dUTP:dTTP se regula de una manera inusual: en todos los tejidos que no serán necesarios en el insecto adulto hay niveles mucho más bajos de la enzima que descompone la dUTP y genera un precursor para la producción de dTTP. Por consiguiente, durante la síntesis del DNA se incorporan a estos tejidos cantidades significativas de uracilo.
Así que durante las etapas larvarias se produce DNA con uracilo que no se va a corregir en los tejidos que van a ser degradados durante la fase de pupa. Como estos insectos carecen de la principal enzima uracil-DNA glicosilasa, en la fase de pupa algunos factores adicionales específicos para DNA con uracilo pueden interpretar este uracilo acumulado como una señal para iniciar la muerte celular. Ya hemos identificado una proteína específica de insectos que parece ser capaz de degradar el DNA con uracilo y estamos investigando si esta enzima se utiliza para iniciar la muerte celular programada.

secuencias de los genes de
anticuerpos provoca una
respuesta de reparación del
DNA, que tiene el efecto de
aumentar la diversidad de los
anticuerpos. Una amplia
reserva de anticuerpos
aumenta la probabilidad de
que el sistema inmune
reconozca a los invasores no
deseados
Imagen cortesía de taramol /
iStockphoto
Errores beneficiosos: el sistema inmune de los vertebrados
También podemos encontrar uracilo en el DNA mucho más cerca: en el sistema inmune de los vertebrados como nosotros. El sistema inmune adaptativo, parte de nuestro sistema inmunológico, produce un gran número de anticuerpos diferentes que están capacitados para protegernos de agentes patógenos específicos. Con el fin de aumentar el número de anticuerpos diferentes que se pueden crear, “mezclamos” la secuencia de DNA en las regiones que codifican para ellos, no solo mediante la recombinación de las secuencias existentes en las células sino también mediante la creación de otras nuevas a través de un aumento enorme de las tasas de mutación, proceso conocido como hipermutación.
La hipermutación se inicia con una enzima específica (una desaminasa inducida por activación) que cambia la citosina por uracilo (véase la Figura 4) en loci concretos del DNA, provocando una respuesta de reparación propensa a errores que el organismo utiliza en su propio beneficio: los “errores” generan nuevas secuencias que se pueden utilizar para hacer anticuerpos diferentes. Sin embargo, este sistema está regulado de forma muy estricta, ya que si se se descontrolara daría lugar a cáncer.
Al considerar todas estas cuestiones necesitamos tener en cuenta el contexto evolutivo. Los organismos vivos han evolucionado en un entorno en continuo cambio, haciendo frente a un cambiante conjunto de desafíos. Por lo tanto, una solución que evite que los errores se incorporen al DNA será ventajosa para la mayoría de los organismos y células, lo que explica por qué el DNA con timina se convirtió en el predominante. Sin embargo, bajo ciertas circunstancias los “errores” pueden ser beneficiosos, por lo que algunas células todavía siguen utilizando uracilo en su DNA.
Resources
- Para saber más sobre el trabajo del grupo de investigación de Beáta Vértessy, consultar: www.enzim.hu/~vertessy
- Para descargar un resumen de la tesis doctoral de Villő Muha, escrita bajo la supervisión de Beáta Vértessy y centrada en DNA con uracilo de Drosophila melanogaster, seguir el enlace: http://teo.elte.hu/minosites/tezis2010_angol/v_muha.pdf
-
La tesis completa está disponible aquí: http://teo.elte.hu/minosites/ertekezes2010/muha_v.pdf
-
Review
Este artículo demuestra que la ciencia nunca duerme, rebatiendo el dogma de que el uracilo solo existe en el RNA. Como explica el artículo, esto no siempre es así. E incluso cuando lo es, ¿por qué es así?
A continuación se muestran ejemplos de preguntas que se pueden utilizar para ayudar a los estudiantes a entender el artículo:
- Describe las estructuras de enlace entre dos pares de bases complementarias en el DNA.
- ¿Cuál de las bases es sustituida en el RNA?
- Describe y dibuja un gráfico de la reparación por vía enzimática que se desencadena cuando se encuentra uracilo en el DNA.
- ¿La proporción de qué moléculas podría ajustarse para impedir que las células cancerosas crezcan y se dividan?
- ¿Por qué es “tolerado” el uracilo en el RNA?
- ¿Qué organismos vivos utilizan uracilo en el DNA y cómo?
Friedlinde Krotscheck, Austria