




L’uracile dans l’AND : erreur ou signal? Understand article
Traduit par Julie Jacq. L’uracile est bien connu comme l’une des bases azotées composant l’ARN, mais on ne le retrouve pas dans l’ADN… ou du moins le croyait-on. Angéla Békési et Beáta G Vértessy se penchent sur le sujet.
Thymine ou uracile?
Notre information génétique est stockée sous forme d’ADN, utilisant quatre bases chimiques que l’on schématise par quatre lettres de l’alphabet. Chaque bloc de construction de l’ADN, ou nucléotide, comporte l’une de ces quatre bases : l’adénine (A), la thymine (T), la cytosine (C) ou la guanine (G; voir Figure 1). Comme l’ont découvert James Watson et Francis Crick, l’ADN prend la forme d’une double hélice, dans laquelle les quatre bases s’associent toujours de la même façon, grâce à des liaisons hydrogènes spécifiques : l’adénine se lie toujours à la thymine, tandis que la guanine se fixe à la cytosine (voir Figures 2 et 3).
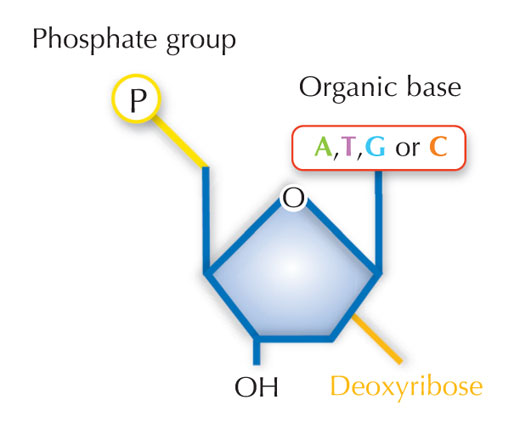
Image reproduite avec l’aimable autorisation de Nicola Graf
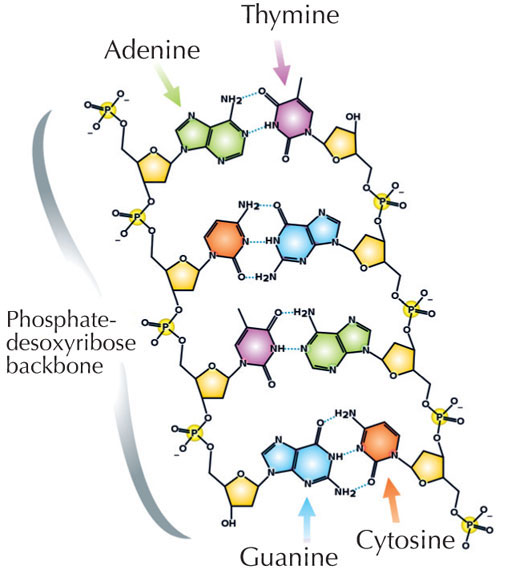
Image reproduite avec l’aimable autorisation de Madeleine Price Ball; Source: Wikimedia Commons
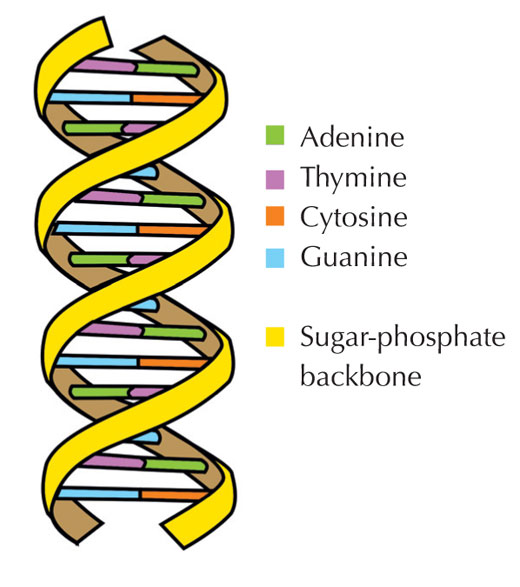
Image reproduite avec l’aimable autorisation de Forluvoft; Source: Wikimedia Commons
Il existe toutefois une cinquième base : l’uracile, qui peut former avec l’adénine les même liaisons hydrogènes que la thymine (voir Figure 4). Mais bien que l’uracile soit normalement présent dans l’ARN, on ne le trouve pas dans l’ADN, où il est remplacé par la thymine. Pourquoi est-ce le cas ?
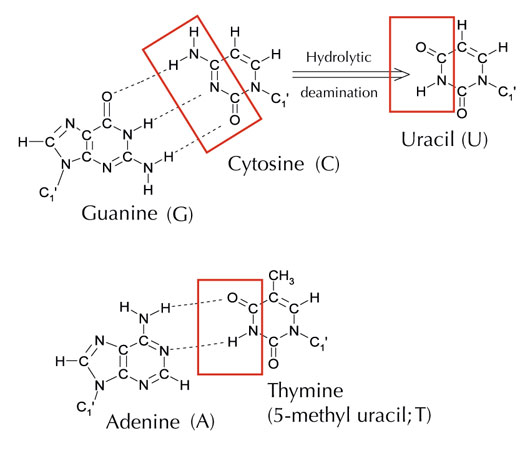
Image reproduite avec l’aimable autorisation de Angéla Békési
Si l’on observe sa composition chimique, on constate que la thymine est en fait une molécule d’uracile possédant un groupe méthyle supplémentaire. Mais quel pourrait être l’avantage, en termes d’évolution, à utiliser cette base à la structure plus complexe dans l’ADN ? Pour le comprendre, il faut s’intéresser à la façon dont les cellules réparent les dommages causé à l’ADN.
hydraulique de la cytosine
peut modifier l’enchaînement
des nucléotides, et par
conséquent altérer les acides
aminés codés par la
séquence d’ADN. Cliquer sur
l’image pour l’agrandir
Image reproduite avec
l’aimable autorisation de Nicola
Graf
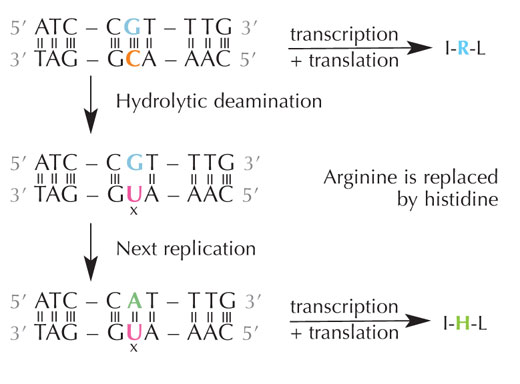
La cytosine peut se transformer spontanément en uracile par un processus connu sous le nom de déamination hydraulique (voir Figure 4). Lorsque cela se produit, la guanine qui était à l’origine liée à cette molécule de cytosine se retrouve en face de l’uracile (rappelons que l’uracile est normalement complémentaire de l’adénine). Par la suite, dès que la cellule commence un nouveau cycle de réplication de l’ADN, cette position face à l’uracile devient occupée par une adénine, plutôt que par la guanine qui se serait trouvée là en temps normal, modifiant donc le code génétique exprimé par cette séquence d’ADN (voir Figure 5). Ce processus de déamination hydraulique est l’un des dommages les plus fréquemment causés à l’ADN, mais il est en général facilement corrigé par la cellule. Quel est donc ce mécanisme de réparation ?
déamination hydraulique.
Cliquer sur l’image pour
l’agrandir
Image reproduite avec
l’aimable autorisation de Nicola
Graf
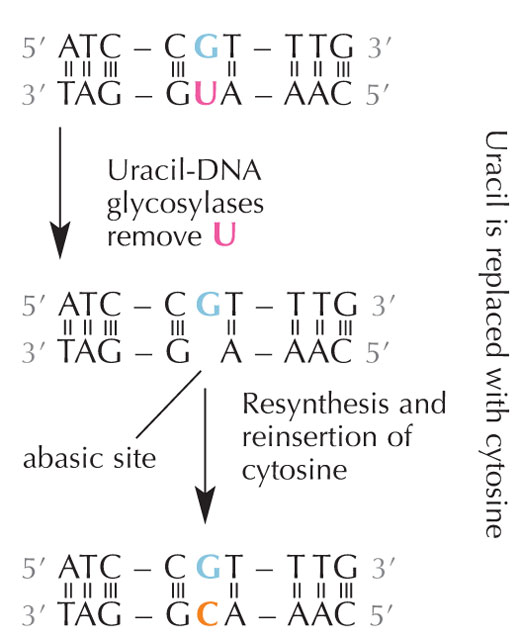
Les cellules possèdent un système de réparation qui peut détecter si un uracile est présent à la place d’une cytosine, et corriger l’erreur avant qu’elle ne soit répliquée et transmise aux cellules-filles. Ce système de réparation est une machinerie complexe utilisant plusieurs enzymes : tout d’abord, une uracile-ADN glycosylase identifie l’uracile dans la séquence d’ADN et l’excise. Ensuite, plusieurs autres enzymes contribuent à l’élimination et à la resynthèse de la partie endommagée de l’ADN, durant laquelle le site abasique (‘vide’) dans le brin d’ADN est complété par une cytosine (voir Figure 6).
Cependant, la forme la plus fréquente d’uracile-ADN glycosylase ne peut pas déterminer quelle est la base à laquelle l’uracile est associé, autrement dit si la présence de l’uracile est normale (lorsqu’il est lié à l’adénine), ou si il s’agit d’une mutation de la cytosine (auquel cas il est lié à la guanine) : en d’autres termes, l’enzyme reconnaît et élimine l’uracile dans les deux cas, qu’il s’agisse ou non d’une mutation. De toute évidence, cela pourrait poser problème. Afin justement d’éviter ce problème, on pense que les cellules auraient développé un mécanisme permettant de lier un groupe méthyle aux molécules d’uraciles ‘correctes’ (c’est à dire associées à de l’adénine), les transformant ainsi en thymine (voir Figure 4). Ce mécanisme permettrait donc à la machinerie cellulaire de reconnaître et exciser un uracile anormal et de réparer l’ADN, sans toucher aux molécules d’uracile liées à un groupe méthyle, celles-ci étant devenues des thymines. (voir Figure 4). Ainsi, au fil du temps, la présence de thymine dans l’ADN est devenue la norme, et la plupart des cellules n’utilisent plus l’uracile que dans l’ARN.
dUTP/dTTP augmente, les
enzymes ADN polymérases
risquent alors d’incorporer
fréquemment de l’uracile à la
place de la thymine lors de la
réplication et de la réparation
de l’ADN. L’uracile-ADN
glycosylase élimine l’uracile
et lance le processus de
réparation de l’ADN, dont
l’une des étapes
intermédiaires est le clivage
du brin d’ADN portant la
lésion. Cependant, il est
possible que l’uracile soit de
nouveau incorporé lors de la
resynthèse de l’ADN, rendant
donc inutile le cycle de
réparation de l’ADN lésé. Au
final, le système de
réparation est submergé, ce
qui conduit à la
fragmentation du
chromosome et à la mort de
la cellule. Cliquer sur l’image
pour l’agrandir
Image reproduite avec
l’aimable autorisation de
Angéla Békési
Pourquoi trouve-t-on toujours de l’uracile dans l’ARN ? L’ARN a une durée de vie beaucoup plus courte que l’ADN et, à de rares exceptions près, son rôle n’est pas de stocker l’information génétique à long terme. Par conséquent, la transformation spontanée de certaines molécules de cytosine en uracile ne représente pas un grand danger pour la cellule si elle se produit dans l’ARN. Il n’y a donc probablement pas eu de pression de sélection poussant la cellule à remplacer l’uracile par de la thymine (plus coûteuse énergiquement à produire car plus complexe) au niveau de l’ARN.
La mort cellulaire par manque de thymine
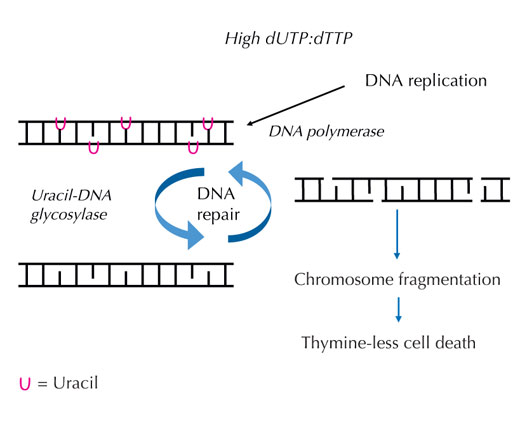
Lors de la synthèse de l’ADN, les enzymes ADN polymérases (qui sont des catalyseurs de la synthèse) ne peuvent pas faire la différence entre la thymine et l’uracile. Elles vérifient seulement que les liaisons hydrogènes sont correctes, autrement dit que les bases complémentaires sont correctement associées entre elles. Mais pour ces enzymes, que l’adénine soit liée à une molécule de thymine ou à une molécule d’uracile n’a pas d’importance. En effet, le taux de désoxyuridine triphosphate (ou dUTP, source de l’uracile) normalement présent dans la cellule est très inférieur au taux de désoxythymidine triphosphate (ou dTTP, source de la thymine), ce qui empêche l’uracile d’être incorporé lors de la synthèse de l’ADN.
Si la régulation de ces deux molécules est perturbée et que la quantité de dUTP par rapport au dTTP augmente, le taux d’uracile incorporé par erreur dans l’ADN augmente également. Le système de réparation (qui, contrairement aux ADN polymérases, sait faire la différence entre uracile et thymine) tente alors d’éliminer l’uracile en activant l’uracile-ADN glycosylase, puis de resynthétiser l’ADN en clivant (coupant) temporairement le brin au niveau du squelette sucre-phosphate. Cependant, si la quantité de dUTP reste encore élevée par rapport au dTTP, cette resynthèse risque d’incorporer encore une fois de l’uracile à la place de la thymine. Ce cycle finit par conduire à la rupture des brins d’ADN et à la fragmentation des chromosomes lorsque ces clivages temporaires de l’ADN deviennent trop nombreux et se produisent trop près les uns des autres (voir Figure 7). Cela entraîne un type particulier de mort cellulaire programmée, appelée mort cellulaire par manque de thymine.
Le processus de mort cellulaire par manque de thymine peut être délibérément exploité pour le traitement du cancer. Les cellules cancéreuses prolifèrent bien plus rapidement que les cellules normales ; par conséquent, elles synthétisent davantage d’ADN pour une période de temps donnée et ont donc besoin de grandes quantités de dUTP. En augmentant le taux de dUTP sans toucher au taux de dTTP, il est donc possible de cibler et éliminer ces cellules cancéreuses.
L’uracile reste présent dans l’ADN
Bien que la plupart des cellules utilisent l’uracile pour la synthèse de l’ARN et la thymine pour celle de l’ADN, il existe quelques exceptions. Chez certains organismes, l’uracile remplace la thymine dans la totalité de l’ADN, tandis que chez d’autres, l’uracile est présent mais seulement dans une partie de l’ADN. Ce phénomène présente-t-il un avantage évolutif pour ces organismes ? Penchons-nous sur quelques exemples.
L’uracile dans l’ADN viral
phage virus infecting a
bacterial cell
Image courtesy of cdascher /
iStockphoto
Deux espèces de phages (virus infectant les bactéries) présentent un ADN dans lequel la thymine a été entièrement remplacée par l’uracile. Nous ignorons encore si ces phages appartiennent à une forme de vie ancienne, n’ayant pas connu les mécanismes d’évolution qui ont conduit à l’incorporation de la thymine dans l’ADN, ou si cette présence d’uracile dans le génome est le résultat d’une nouvelle stratégie évolutive. Nous ignorons également pourquoi ces phages utilisent de l’uracile à la place de la thymine, mais il est possible que ce choix joue un rôle essentiel dans le cycle de vie de ces virus. Si c’est bien le cas, il serait alors normal que ces virus s’assurent que l’uracile présent dans leur ADN ne soit pas remplacé par de la thymine. Et en effet, il a été prouvé que l’un de ces phages possède un gène inhibant l’uracile-ADN glycosylase de la cellule hôte, empêchant donc le génome viral d’être ‘réparé’ par les enzymes de l’hôte.
La mort cellulaire programmée dans le cycle de vie de l’insecte

les fourmis ne possèdent pas
l’enzyme capable d’éliminer
l’uracile de leur ADN
Image reproduite avec
l’aimable autorisation de
spxChrome / iStockphoto and
Nicola Graf
L’uracile incorporé dans l’ADN semble également jouer un rôle dans le développement des endoptérygotes (insectes qui connaissent une métamorphose au cours de leur vie : les fourmis et les papillons en font partie, mais pas les sauterelles ou les termites). Ces insectes ne possèdent pas le gène principal codant pour l’uracile-ADN glycosylase, l’enzyme normalement chargée d’éliminer l’uracile de l’ADN.
De plus, notre propre recherche a montré que, chez la larve de la mouche drosophile Drosophila melanogaster, le rapport entre dUTP et dTTP fait l’objet d’une régulation inhabituelle : au niveau de tous les tissus qui disparaissent chez l’insecte adulte, on trouve des quantités beaucoup plus basses de l’enzyme chargée de dégrader le dUTP et de générer un précurseur pour la production de dTTP. Par conséquent, des quantités non-négligeables d’uracile sont incorporées dans ces tissus lors de la synthèse de l’ADN.
Il apparaît donc que, durant le stade larvaire, l’uracile est incorporé dans l’ADN sans être corrigé au niveau des tissus destinés à être dégradés lors du stade nymphal. Etant donné que ces insectes ne possèdent pas la principale enzyme uracile-ADN glycosylase, il est possible que, lors du stade nymphal, des facteurs supplémentaires spécifiques à l’ADN porteur d’uracile puissent reconnaître cette accumulation d’uracile comme un signal commandant à l’organisme de déclencher la mort cellulaire. Nous sommes déjà parvenus à identifier une protéine spécifique à l’insecte qui semble capable de dégrader l’ADN porteur d’uracile, et nous cherchons actuellement à déterminer si cette enzyme est utilisée pour déclencher la mort cellulaire programmée.

niveau des séquences de
gènes codant pour les
anticorps déclenche un cycle
de réparation de l’ADN, ce qui
a pour effet d’accroître la
diversité des protéines
anticorps. Un pool
d’anticorps important signifie
que le système immunitaire
aura une plus grande chance
d’identifier les envahisseurs
de l’organisme
Image reproduite avec
l’aimable autorisation de
taramol / iStockphoto
Les erreurs bénéfiques : le système immunitaire des vertébrés
On retrouve toutefois de l’uracile présent dans l’ADN chez des espèces plus proches de la nôtre, et plus précisément au niveau du système immunitaire des vertébrés. Une partie de notre système immunitaire, et plus exactement le système immunitaire acquis, produit de grandes quantités de divers anticorps qui ont pour but de nous protéger d’agents pathogènes spécifiques. Afin d’augmenter le nombre d’anticorps différents pouvant être produits par notre organisme, nous réordonnons la séquence d’ADN dans les régions codant pour ces anticorps, soit par recombinaison dans les cellules de séquences déjà existantes, soit en créant de nouvelles séquences par hypermutation (c’est à dire en augmentant la fréquence d’apparition de mutations dans l’ADN).
L’hypermutation commence par l’action d’une enzyme spécifique (une déaminase induite par activation) qui transforme la cytosine en uracile (voir Figure 4) au niveau d’un loci d’ADN spécifique. Cela entraîne des erreurs lors de la réparation de l’ADN, mais ces ‘erreurs’ peuvent être exploitées par l’organisme car elles génèrent de nouvelles séquences d’ADN pouvant être utilisées pour produire de nouveaux anticorps. Cependant, ce système fait l’objet d’une régulation extrêmement stricte, car il pourrait générer des cancers si l’organisme venait à en perdre le contrôle.
Pour mieux comprendre pourquoi l’organisme choisit d’utiliser la thymine ou l’uracile, il faut prendre en compte le contexte évolutif. Les organismes vivants ont évolué dans un environnement qui change constamment, et pose donc en permanence de nouveaux défis. Par conséquent, la plupart des cellules et des organismes trouvent avantageux d’avoir un moyen d’empêcher les erreurs d’être incorporées dans l’ADN, ce qui explique pourquoi l’utilisation de la thymine lors de la synthèse est devenue la norme. Il existe toutefois certaines circonstances dans lesquelles ces mêmes ‘erreurs’ peuvent être bénéfiques. C’est la raison pour laquelle certaines cellules incorporent encore l’uracile dans leur ADN.
Resources
- Pour en savoir plus sur le travail de recherche de l’équipe de Beáta Vértessy, rendez-vous sur :www.enzim.hu/~vertessy
- Pour télécharger un résumé de la thèse de Villő Muha, rédigée sous la supervision de Beáta Vértessy et portant sur la présence de l’uracile dans l’ADN de la Drosophila melanogaster, cliquez sur ce lien : http://teo.elte.hu/minosites/tezis2010_angol/v_muha.pdf
-
La thèse complète est disponible sur : http://teo.elte.hu/minosites/ertekezes2010/muha_v.pdf
-
Review
On n’arrête pas la science. Cet article en est la preuve, allant à l’encontre des convictions en prouvant que l’uracile peut se rencontrer ailleurs que dans l’ARN. Il démontre également que lorsque la présence de l’uracile est limitée à l’ARN, ce n’est pas sans raison.
Pour aider les étudiants à comprendre cet article, voici quelques questions que vous pourriez poser:
- Décrivez les liaisons existant entre les deux paires de bases complémentaires de l’ADN.
- Quelle base est remplacée dans l’ARN?
- Décrivez et dessinez un graphique montrant l’action de l’enzyme de réparation lorsque la présence d’uracile est détectée dans l’ADN.
- Afin de stopper la division et la prolifération des cellules cancéreuses, il est possible de modifier le ratio entre deux molécules. Lesquelles ?
- Pourquoi l’organisme ‘tolère’-t-il l’uracile dans l’ARN ?
- Quels organismes incorporent de l’uracile dans l’ADN et pourquoi?
Friedlinde Krotscheck, Autriche