




La sélection naturelle vue au niveau moléculaire Understand article
Traduit par Maurice A. Casimir. Nous savons que des séquences génétiques particulières peuvent nous aider à survivre dans notre environnement – c’est la base de l’évolution. Mais démontrer quelles séquences génétiques sont en notre faveur et la manière dont elles nous aident à…

iStockphoto
Lorsque les humains quittèrent pour la première fois l’Afrique il y a 150.000 ans, pour s’installer dans les vallées du Tigre et de l’Euphrate, faire voile vers les îles indonésiennes et franchir le Détroit de Behring vers l’Amérique, ils durent faire face à de nombreux défis. Venant de savanes africaines chaudes et desséchées, les populations durent s’adapter aux conditions locales, et leur physiologie et leur apparence se modifièrent en conséquence au fil des générations (Harris & Meyer, 2008). Leur peau devint plus pâle après qu’ils eurent vécu dans des régions moins ensoleillées (Lamason et al., 2005). Les populations dont les membres buvaient du lait d’animaux domestiqués conservèrent la capacité de digérer le lactose à l’âge adulte, une caractéristique perdue dès la fin de l’enfance parmi les groupes à régime non lacté (Tishkoff et al., 2007). Les populations consommant une nourriture riche en amidon produisirent plus d’amylase salivaire, l’enzyme qui aide à décomposer l’amidon (Perry et al., 2007).
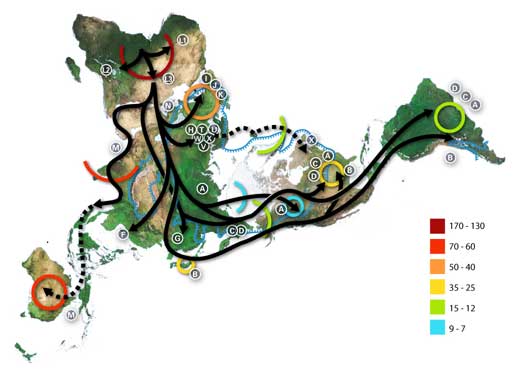
Les nombres représentent des milliers d’années avant l’époque actuelle.
La ligne bleue représente la zone recouverte de glace ou de toundra durant la dernière grande glaciation. Les lettres correspondent à des haplogroupes d’ADN mitochondrial (lignages maternels purs). On peut utiliser les haplogroupes pour définir des populations génétiques et ils sont souvent localisés géographiquement.
Par exemple, on a communément les répartitions suivantes pour les haplogroupes de mtDNA:
Afrique: L, L1, L2, L3, L3
Proche Orient: J, N
Europe du Sud: J, K
Europe entière: H, V
Europe du Nord: T, U, X
Asie: A, B, C, D, E, F, G (nota: M est composé de C, D, E et G)
Américains précolombiens: A, B, C, D et parfois X.
Cliquer sur l’image pour l’agrandir
Illustration reproduite avec l’aimable autorisation d’Avsa; source: Wikimedia Commons
L’on pense qu’au moins certains de ces changements ont été la conséquence d’une sélection positive (voir glossaire pour les termes en italique). Ceci implique que, dans le passé et dans un environnement particulier (la pression de sélection), les organismes qui se trouvèrent avoir une séquence ADN avantageuse survécurent et laissèrent plus de descendants que ceux dotés d’une séquence différente, moins en leur faveur. De nos jours, en utilisant les séquences génomiques de nombreuses espèces, dont les humains et les êtres vivants les plus proches dans l’arbre de l’évolution, les scientifiques peuvent comparer des caractères hérités et des séquences d’ADN de populations ou espèces aux modes de vie différents et dans des environnements différents pour identifier les séquences ayant pu jouer un rôle dans les adaptations. Ceci permet à leur tour aux chercheurs d’étudier la fonction d’une séquence d’ADN et sa valeur d’adaptation potentielle pour un organisme.

l’aimable autorisation de
JBryson / iStockphoto
Certains des gènes connus pour affecter la couleur de la peau chez les humains montrent une localisation géographique spécifique dans la variation des séquences; en particulier, des comparaisons de séquences entre les populations européennes et africaines suggèrent que des variations de couleur de peau sont dues à la sélection positive. Une couleur de peau claire est corrélée positivement avec des latitudes croissantes, et l’on a proposé plusieurs hypothèses pour expliquer ses effets potentiellement avantageux.Une hypothèse, qui suppose qu’une peau claire favorise la production de vitamine D, est renforcée par des observations selon lesquelles les peuples à la peau sombre vivant sous des latitudes élevées souffrent d’une carence en vitaline D. En outre, une peau claire est plus sensible aux effets néfastes de l’ensoleillement: une exposition accrue au soleil est en corrélation avec une occurrence plus grande de cancer de la peau chez les peuples à peau claire. Par conséquent, la pâleur de la peau chez les populations humaines vivant sous des latitudes élevées peut constituer un compromis au niveau de l’évolution entre la protection contre les effets cancérigènes de l’ensoleillement et la capacité d’une production suffisante d’une vitamine essentielle.
électronique à balayage d’une
cellule atteinte de
drépanocytose et d’autres cellules sanguines
Illustration reproduite avec
l’aimable autorisation de l’EM
Unit, UCL Medical School, Royal Free Campus / Wellcome Images
Bien qu’il s’agisse d’une hypothèse solide, elle ne s’appuie que sur des preuves indirectes. Une démonstration directe de la valeur d’adaptation de ce trait nécessiterait de mesurer si, sous des latitudes plus élevées, les individus à la peau plus pâle montrent un niveau de survie et de reproduction accru. De telles démonstrations sont cependant difficiles pour nos espèces: des expériences de survie (dans lesquelles des organismes avec des traits différents sont exposés à un environnement donné pour voir lesquels survivent) ne peuvent être réalisées sur l’être humain, et la longue période s’écoulant entre deux générations rend difficile la recherche de différences entre les taux de reproduction. Les circonstances pour lesquelles il est possible d’observer la valeur d’adaptation d’un trait donné chez les humains sont donc très limitées – mais elles existent effectivement.
gorge de sang. Cette espèce,
Anopheles stephensi, est le
vecteur de transmission de la
malaria en India et au
Pakistan
Illustration reproduite avec
l’aimable autorisation de Hugh
Sturrock / Wellcome Images
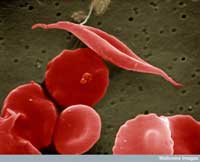
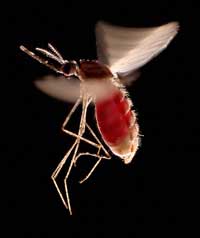
Un exemple concerne deux maladies: la drépanocytose et la malaria. Le gène impliqué dans la drépanocytose a deux variantes ou allèles: un allèle ‘normal’ et un allèle dit S. Les personnes avec deux allèles S souffrent d’une drépanocytose sérieuse, alors que celles avec un allèle S et un allèle normal ne présentent pas de symptomes aussi sévères. Les données sur la mortalité suggèrent que l’allèle S peut cependant offrir un avantage: chez les populations exposées au parasite de la malaria, les personnes présentant un allèle S et un allèle normal vont plus probablement survivre que celles portant deux allèles normaux, car le parasite (Plasmodium falciparum) a besoin de cellules sanguines en bonne santé pour envahir l’organisme et s’y multiplier. La fréquence de l’allèle causant la drépanocytose augmente donc dans les groupes exposés à la malaria – l’allèle s’adapte à son environnement.
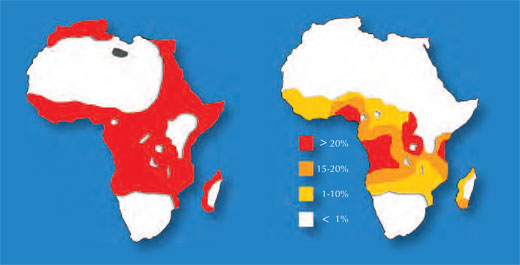
Illustration reproduite avec l’aimable autorisation d’Anthony Allison
Un autre exemple montrant la valeur d’adaptation d’un trait humain concerne un fragment du chromosome 17, dont l’on sait qu’il a été inverti chez nos ancêtres il y a plus de trois millions d’années (Stefansson et al., 2005). Que cette variante se soit disséminée parmi les populations européennes suggère qu’elle a fait l’objet d’une sélection positive – elle a conféré un avantage à ceux qui la portent. En dressant le génotype de près de 30.000 Islandais, des scientifiques étudiant cette hypothèse ont pu déterminer que, dans les 80 dernières années, les insulaires porteurs de la séquence avaient en moyenne 3.2% de descendants par génération que ceux dotés de la séquence normale, une explication plausible de la manière dont la variante en est est venue à se répandre si rapidement.
« Bien que les deux exemples montrent clairement l’action de la sélection positive sur les êtres humains, l’on ne comprend pas bien les mécanismes moléculaires conférant leurs avantages aux variations de séquences et il faut les rechercher au cas par cas. Pour élucider les relations causales entre des séquences d’ADN potentiellement adaptatives et leur valeur sélective au niveau individuel, les scientifiques se tournent vers des organismes sur lequels il est plus facile d’expérimenter que sur des êtres humains.
Par exemple, la couleur du pelage de la souris d’Oldfield, Peromyscus polionotus, imite celle du sol de l’habitat, ce qui lui fournit un camouflage. Les souris vivant dans les sables blancs des plages de Floride sont beaucoup plus claires que les souris de l’intérieur des terres de la même espèce. La valeur d’adaptation de ce trait a été démontrée il y a plus de 30 ans: les souris avec un pelage imitant la couleur du sol étaient dévorées moins fréquemment par les chouettes que les autres souris moins bien camouflées. Cependant, les scientifiques n’ont identifié que récemment les locus génétiques dans lesquels se cachait ce trait d’adaptation (Hoekstra et al., 2006): la variation dans la couleur du pelage dépend largement de différents allèles du gène McR1. La protéine encodée par ce gène agit comme un commutateur biochimique entraînant la production soit d’eumélanine (tyrosine), un pigment foncé de la peau, soit de phéomélanine (cystéine), un pigment clair. Les différents allèles du gène McR1 activent le système de production du pigment de façon différenciée, favorisant la production d’un pigment ou de l’autre.
électronique à balayage de
groupes de bactéries de
Staphylococcus aureus
résistantes à la méticilline
Illustration reproduite avec
l’aimable autorisation d’Annie
Cavanagh / Wellcome Images
Un autre exemple de relation causale démontrée implique Staphylococcus aureus, une bactérie pouvant être responsable de diverses maladies dont la pneumonie ou l’inflammation de valvule cardiaque. Dans une expérience in vivo rare, un patient victime d’infections récurrentes dues au S.aureus a été traité pendant trois mois à la vancomycine, l’un des quelques antibiotiques encore efficaces contre la bactérie. Avant le traitement et à intervalles réguliers durant celui-ci, les scientifiques collectèrent des échantillons (des isolats) du germe pathogène et en séquencèrent le génome entier dans les premier et dernier isolats. Lorsqu’ils comparèrent les trois millions de paires de base (les ‘lettres’ du code génétique) constituant l’ADN de cette bactérie, ils ne trouvèrent que 35 différences entre les premier et dernier isolats.
En séquençant partiellement les isolats intermédiaires, les scientifiques déterminèrent l’ordre dans lequel ces changements ont dû se produire. En testant la résistance de la bactérie à la vancomycine in vitro dans les différents isolats, ils purent corréler les changements génétiques particuliers ayant des effets sur la croissance de la bactérie et la réponse au médicament. Par exemple, les bactéries des premier et second isolats différaient par six substitutions de nucléotide (changements de ‘lettres’) dans deux gènes. Ces six mutations étaient clairement avantageuses en elles-mêmes: elles multipliaient la tolérance de la bactérie à la vancolycine par un facteur de quatre, ce qui permettait à la bactérie de mener à bien ces mutations pour survivre et mieux se reproduire, en devenant plus commune dans le corps du patient. Vingt six mutations successives dans les semaines suivantes du traitement doublèrent la tolérance, produisant effectivement une lignée de S.aureus tolérant la vancomycine (Mwangi et al., 2007).
En bref, étudier les bases moléculaires de l’évolution par adaptation dans des populations sauvages n’est pas aisé. Parmi les défis à relever, citons la définition des pressions sélectives, l’identification des séquences d’ADN se cachant derrière les caractères hérités associés, la mesure de l’adaptation à la fonction, la découverte des explications mécanistes sur la manière dont les changements de séquence modifient les caractères hérités par l’adaptation. Cependant, ces études sont aujourd’hui devenues réalisables grâce à l’utilisation d’organismes modèles et à de récents développements technologiques, ce qui augmente notre compréhension de la manière dont des changements spécifiques au niveau génétique permettent à des organismes de s’adapter à leur environnement.
Glossaire
Valeur d’adaptation: un caractère hérité a une valeur d’adaptation s’il permet à un organisme individuel de mieux survivre et de se reproduire dans un environnement donné que ceux qui ne possèdent pas ce caractère. Plus formellement, un caractère hérité est considéré adaptatif s’il augmente la valeur sélective.
Allèle: une variante d’un gène.
Valeur sélective: un terme de biologie de l’évolution et de génétique des populations formellement difficile à définir; il décrit le nombre moyen de descendants sur une génération associé à un génotype comparé à un autre génotype d’une population. Ainsi, les génotypes produisant plus de descendants ont une valeur sélective supérieure. Pour une bonne vue d’ensemble de la valeur sélective et du génotype, voir Wikipediaw1.
Génome: l’ADN total d’un organisme. On comprend généralement sous ce terme l’ADN nucléaire, par opposition à l’ADN mitochondrial ou plaste. Pour de plus amples informations, voir ‘Qu’est-ce qu’un génome?’ sur le site Internet de la Bibliothèque Nationale de Médecine des Etats Unisw2.
Sélection positive: la selection naturelle est l’un des mécanismes de l’évolution; elle décrit la survie et la reproduction différenciées des organismes individuels dans un environnement donné. La sélection naturelle est dite ‘positive’ lorsqu’elle met en avant certains caractères hérités aidant les organismes individuels en disposant de survivre et de se reproduire mieux que le sautres.
Pression de selection: une caractéristique de l’environnement (comme température; présence de parasites; prédation ou agression par des membres de la même espèce) qui impose une survie et une reproduction différenciées des organismes individuels.
Caractère hérité: une particularité des caractéristiques d’un organisme ou un ensemble de particularités de celui-ci (comme taille; résistance aux antibiotiques, capacité de reconnaître les couleurs ou d’enrouler sa langue).
Acknowledgements
The author is grateful to David Hughes, Mehmet Somel and Ania Lorenc for helpful comments on the article.
References
- Harris EE, Meyer D (2006) The molecular signature of selection underlying human adaptations. American Journal of Physical Anthropology 131(S43): 89-130. doi: 10.1002/ajpa.20518
- Cet article fournit une bonne vue d’ensemble de la recherche sur l’évolution au niveau moléculaire chez les humains.
- Hoekstra H et al. (2006) A single amino acid mutation contributes to adaptive beach mouse color pattern. Science 313: 101-104. doi: 10.1126/science.1126121
- L’article ci-dessus et d’autres publications sur la coloration du pelage de la souris par le groupe de recherche de Hopi Hoekstra sont disponibles sur le site Internet de l’Université Harvard. Voir: www.oeb.harvard.edu/faculty/hoekstra/Links/PublicationsPage.html
- Voir également la publication ultérieure dans laquelle est décrite la découverte d’Agouti, un régulateur négatif de McR1 contribuant à l’adaptation de la couleur du pelage chez Peromyscus:
- Steiner CC, Weber JN, Hoekstra HE (2007) Adaptive variation in beach mice produced by two interacting pigmentation genes. PLoS Biology 5: e219. doi: 0.1371/journal.pbio.0050219
- Cet article ainsi que tous les autres articles de PLoS Biology sont disponibles gratuitement en ligne.
- Les articles suivants passent en revue l’adaptation de la pigmentation chez les vertébrés:
- Hoekstra HE (2006) Genetics, development and evolution of adaptive pigmentation in vertebrates. Heredity 97: 222-234. doi: 10.1038/sj.hdy.6800861
- Cet article peut être téléchargé gratuitement sur le site Internet du journal Heredity: www.nature.com/hdy
- Une vue d’ensemble des plus récentes recherches de Hoekstra est disponible sur le blog de John Hawks: http://johnhawks.net/weblog/topics/evolution/selection/hoekstra-2009-adaptive-pigmentation.html
- Lamason RL et al. (2005) SLC24A5, a putative cation exchanger, affects pigmentation in zebrafish and humans. Science 310: 1782-1786. doi: 10.1126/science.1116238
- Mwangi MM et al. (2007) Tracking the in vivo evolution of multidrug resistance in Staphylococcus aureus by whole-genome sequencing. Proceedings of the National Academy of Sciences of the United States of America 104: 9451-9456. doi: 10.1073/pnas.0609839104
- Perry GH et al. (2007) Diet and the evolution of human amylase gene copy number variation. Nature Genetics 39: 1256-1260. doi: 10.1038/ng2123
- Voir également une vue d’ensemble de cette recherche à Panda’s Thumb: http://pandasthumb.org/archives/2008/12/amylase-and-hum.html
- Stefansson H et al. (2005) A common inversion under selection in Europeans. Nature Genetics 37: 129-137. doi: 10.1038/ng1508
- Voir également une vue d’ensemble de l’article à Evolgen: http://evolgen.blogspot.com/2005/02/human-inversion-under-selection.html
- Tishkoff SA et al. (2006) Convergent adaptation of human lactase persistence in Africa and Europe. Nature Genetics 39: 31-40. doi: 10.1038/ng1946
- Voir également une vue d’ensemble de cette recherche dans The New York Times: www.nytimes.com/2006/12/10/science/10cnd-evolve.html?_r=1
Web References
- w1 – Pour une bonne vue d’ensemble des termes ‘valeur sélective’ et ‘génotype’, voir Wikipedia: http://en.wikipedia.org/wiki/Fitness_(biology) and http://en.wikipedia.org/wiki/Genotype
- w2 – Pour de plus amples informations sur les génomes et le Human Genome Project, voir ‘Qu’est-ce qu’un génome?’ sur le site Internet de la Bibliothèque Nationale d eMédecine des Etats Unis: http://ghr.nlm.nih.gov/handbook/hgp/genome
Resources
- Si vous avez trouvé cet article intéressant, vous pouvez avoir envie de lire certains des articles de Science in School sur l’évolution:
- Haubold B (2010) Review of Why Evolution is True. Science in School 14. www.scienceinschool.org/2010/issue14/evotrue
- Leigh V (2008) Interview de Steve Jones: La menace du créationnisme. Science in School 9. www.scienceinschool.org/2008/issue9/stevejones/french
- Patterson L (2010) Aller plus avant dans l’évolution avec l’amphioxus. Science in School 14. www.scienceinschool.org/2010/issue14/amphioxus/french
- Pongsophon P, Roadrangka V and Campbell A (2007) Counting Buttons: demonstrating the Hardy-Weinberg principle. Science in School 6: 30-35. www.scienceinschool.org/2007/issue6/hardyweinberg
- Pour plus d’informations sur la malaria, voir:
- Hodge R (2006) Fighting malaria on a new front. Science in School 1: 72-75. www.scienceinschool.org/2006/issue1/malaria
- Pour en apprendre advantage sur la structure de l’amidon, que l’amylase salivaire aide à décomposer, voir:
- Cornuéjols D (2010) L’amidon: une structure mystérieuse. Science in School 14. www.scienceinschool.org/2010/issue14/starch/french
Review
L’article décrit un éventail d’exemples intéressants des adaptations de l’évolution au niveau moléculaire chez les humains. La difficulté à trouver des relations causales entre les séquences d’ADN adaptatives et la valeur sélective individuelle chez les humains et le besoin d’utiliser d’autres organismes pour l’expérimentation sont mis en lumière.
L’article fournit un excellent matériau pour des questions centrées sur la compréhension de la sélection naturelle et la valeur sélective chez les humains et des organismes expérimentaux. Par exemple:
- Expliquez les processus impliqués dans la sélection naturelle.
- Qu’entendez-vous par ‘valeur sélective’?
- Expliquer comment l’allèle S confère un avantage sélectif chez quelques populations humaines.
- Quels sont les problèmes rencontrés dans l’établissement de relations causales entre les séquences d’ADN adaptatives et la valeur sélective chez les humains?
- Elaborer un organigramme expliquant la valeur d’adaptation de la couleur du pelage chez la souris d’Oldfield.
- Comment les scientifiques ont-ils pu corréler les modifications génétiques chez Staphylococcus aureus avec la croissance bactérienne et la réaction à l’antibiotique?
Cet article permet également aux élèves de rechercher le lien entre l’ADN, la séquence d’aminoacides, la structure protéinique et la leur fonction dans la drépanocytose. Le texte est adapté à la conduite d’une discussion en classe sur les méthodes et les problèmes associés à l’étude de la base moléculaire des essais génétiques dans des populations humaines. Des études interdisciplinaires pourraient être organisées autour de l’histoire des sciences et la génétique des populations du point de vue de l’évolution.
Mary Brenan, Royaume-Uni