




Synchrotron light illuminates the orang-utan’s obscure origins Understand article
Paul Tafforeau from the University of Poitiers and the European Synchrotron Radiation Facility in Grenoble, France, explains what synchrotron X-ray studies of fossil teeth can tell us about the evolution of orang-utans – and our own origins.

Today, the hominoids (apes) are represented by only five genera: gibbons, orang-utans, gorillas, chimpanzees and our own species, Homo sapiens. In the Miocene period, between 20 and 6 million years (MY) ago, however, this was a much more diverse group, as demonstrated by the numerous (approximately 20) fossil genera that have been discovered in Africa, Asia and Europe.
The most ancient hominoids lived around 20 million MY ago in Africa. Outside Africa, the first hominoids date from around 16.5 MY ago, but many different forms are known to have lived in Europe and Asia 12-6 MY ago. Subsequently, the group declined in diversity, probably due to climate changes. Several hypotheses have been suggested to explain the phylogenetic (evolutionary) relationships of the fossil and extant (modern) species and to describe their paleobiogeography – that is, their geographic migration and diversification.
The traditional hypothesis is that Africa was the principal area of diversification during the whole Miocene period, with successive migrations to Europe and Asia followed by local evolution on all three continents (map A).
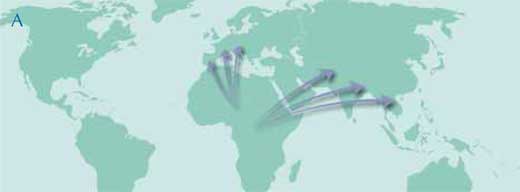
Image courtesy of Petra Riedinger
Over the last few years, however, a new hypothesis has emerged. It also proposes that the hominoids originated in Africa, but that primitive forms then migrated towards Asia, and after a progressive extinction in Africa, Africa and Europe were repopulated by successive migrations from Asia (map B). According to this hypothesis, the principal area of hominoid diversification was Asia instead of Africa. The key fossils to test these two scenarios should therefore be found in Asia.
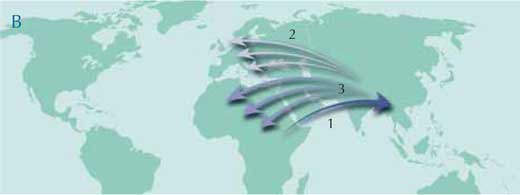
Image courtesy of Petra Riedinger
In 2003, research in Thailand led to the discovery of a previously unknown species of fossil hominoid from approximately 12 MY ago (Chaimanee et al., 2003). About 20 isolated teeth (figure 1A), attributed to several male and female individuals, showed that this species was a large hominoid with a strong sexual dimorphism – the male was much larger than the female, with more developed canines. It was named cf. Lufengpithecus chiangmuanensis.
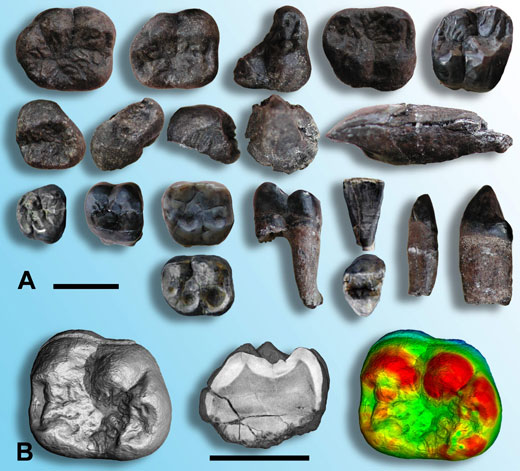
Image courtesy of Paul Tafforeau
Dental enamel is the most highly mineralised tissue found in vertebrates; consequently, teeth are very strong and resistant to mechanical or chemical erosion. This, together with the fact that they contain very little organic matter to be decomposed by micro-organisms, means that teeth are often better preserved during the fossilisation process than the rest of the organism, including bones. Indeed, the vast majority of the vertebrate fossil record consists of teeth and most palaeontological (fossil) studies on vertebrate remains (including hominoids) are based on teeth. Fortunately for the palaeontologist, teeth have many anatomical characteristics linked to both phylogeny and function, whether for feeding or as sexual attributes (the males of many primates have particularly well-developed canines, for example).
In addition to a traditional palaeontological study of external dental morphology, the internal structure of the teeth of cf. L. chiangmuanensis was studied in a non-destructive way, providing results of previously unattainable quality (Chaimanee et al., 2003; Tafforeau, 2004; Tafforeau et al., 2006). At the European Synchrotron Radiation Facility (ESRF)w1 in Grenoble, France, the teeth were imaged in three dimensions using X-ray synchrotron microtomography. This made it possible to quantify precisely the thickness and the distribution of enamel (figure 1B) without having to cut the teeth open. The combination of traditional palaeontology and microtomographic analyses revealed that this fossil species is more similar to modern orang-utans than to any other known fossil hominoids in Asia. It was therefore proposed that cf. L. chiangmuanensis could be an ancestor of the orang-utans.
In 2004, a further fossil, approximately 7 MY old, was discovered in Thailand. This beautiful mandible (lower jawbone; figure 2A) was assigned to yet another new species: Khoratpithecus piriyai (Chaimanee et al., 2004). On the basis of its overall size, the size of the canines and bone morphology, the mandible was thought to have come from a male. A subsequent comparison of this fossil with thecf. L. chiangmuanensis teeth showed that these two forms were more similar and therefore more closely related to one another than to any other fossil or extant species. The older species, cf. L. chiangmuanensis, was then reattributed the new genus and renamed Khoratpithecus chiangmuanensis.
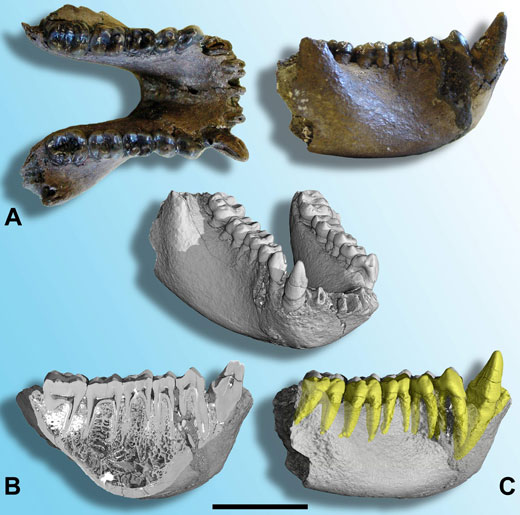
Image courtesy of Paul Tafforeau
K. piriyai’s mandible was also imaged at the ESRF using X-ray synchrotron microtomography, resulting in the first high-quality scan of such a large fossil of a hominoid. Using this exceptional data set, it was possible to analyse the dental structure and the bone architecture (figure 2B), and to virtually extract the teeth from the right side of the mandible to study the size and the shape of tooth roots (figure 2C; Tafforeau, 2004; Tafforeau et al., 2006; Chaimanee et al., 2006).
The 3D data of K. chiangmuanensis’s teeth were used in a virtual reconstruction of the jaws (figure 3), which allowed further comparisons of the general morphology of the jaws and the relative sizes of teeth in the two species of Khoratpithecus. This showed that their similarities were even stronger than initially thought and fully justified their classification in a single genus, which had been made on the basis of traditional palaeontological studies.
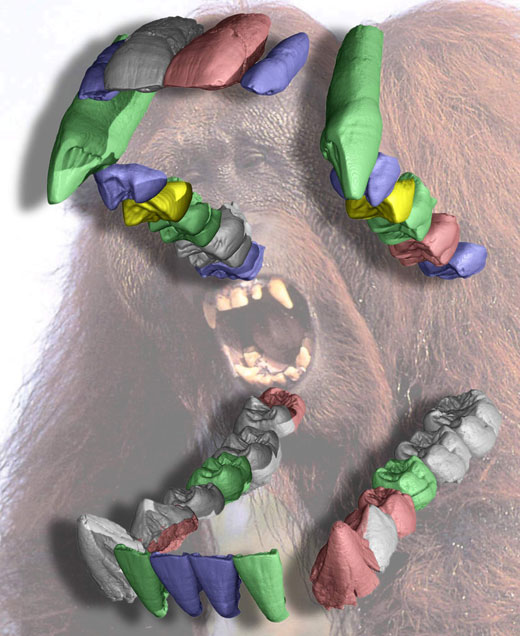
Image courtesy of Paul Tafforeau
Using synchrotron X-ray imaging of these fossils, it was possible to reveal anatomical characteristics that would otherwise have been impossible to study without destroying the fossils. These cutting-edge analyses, combined with a more traditional palaeontological study, revealed that Khorapithecus is the known genus most closely related to the orang-utans. Nevertheless, the studies also showed that although K. piriyai (the more recent species) had many anatomical features that are typical of the orang-utans, it also displayed specialised features. Among them, K. piriyai had a canine morphology very unlike the modern orang-utans, with a dagger-like canine and an extremely enlarged third molar. From what is known from other lineages, it is unlikely that so many specialisations would have evolved and subsequently disappeared.
Therefore, although related, K. piriyai is almost certainly not a direct ancestor of the modern orang-utans. K. chiangmuanensis, the older species with fewer specialised dental characteristics, is more likely to be a possible ancestor of the orang-utans. Although new fossils are necessary to test the hypothesis, it is possible that the orang-utan’s lineage was derived from primitive forms of the Khoratpithecus genus and that the two branches evolved in different ways, giving rise both to the orang-utans and to later species of Khoratpithecus, such as K. piriyai.
We can now return to our two hypotheses of hominoid evolution: African and Asian diversification. The two Khoratpithecus species, as well as the numerous other fossil hominoid taxa from Asia, show a higher diversity than African hominoid fossils during the same period. Moreover, both very specialised and primitive species have been found in Asia. Together, the high levels of diversity and the wide geographic distribution in geological strata of Asian hominoids (from 16.5 MY ago to the present) strongly suggest that Asia was an important, perhaps even the principal, centre of diversification for Miocene hominoids. Increasingly, therefore, modern palaeological research is lending weight to the newer hypothesis of hominoid evolution: our hominoid ancestors originated in Africa, after which they diversified in Asia and repopulated Africa and Europe.
References
- Chaimanee Y et al. (2003) A Middle Miocene hominoid from Thailand and orangutan origins. Nature 422: 61-65. doi:10.1038/nature01449.
- Download the article free of charge here, or subscribe to Nature today: www.nature.com/subscribe
- Chaimanee Y et al. (2004) A new orang-utan relative from the Late Miocene of Thailand. Nature 427: 439-441. doi:10.1038/nature02245.
- Download the article free of charge here, or subscribe to Nature today: www.nature.com/subscribe
- Chaimanee Y et al. (2006) Khoratpithecus piriyai, a late Miocene hominoid of Thailand. American Journal of Physical Anthropology 131: 311-323. doi:10.1002/ajpa.20437
- Tafforeau P (2004) Aspects phylogénétiques et fonctionnels de la microstructure de l’émail dentaire et de la structure tridimensionnelle des molaires chez les primates fossiles et actuels : apports de la microtomographie à rayonnement X synchrotron. Ph.D. Thesis, Université de Montpellier II, France
- Tafforeau P et al. (2006) Applications of X-ray synchrotron microtomography for non-destructive 3D studies of paleontological specimens. Applied Physics A, Materials Science & Processing 83: 195-202. doi:10.1007/s00339-006-3507-2
Web References
- w1 – The European Synchrotron Radiation Facility (ESRF) is an international facility that operates, maintains and develops the most powerful synchrotron light source in Europe, with 18 participating countries. More than 5000 researchers come to the ESRF every year to use the light source and its associated instruments.
- ESRF is a member of EIROforum, a collaboration of seven European inter-governmental research organisations, and the publishers of Science in School.
Resources
- An explanation of the use of synchrotron light at ESRF is available here.
- Tafforeau P et al. (2005) Synchrotron Radiation Microtomography: A Tool for Paleontology. ESRF Newsletter 42: 22-23.
- For information about the University of Poitiers, see: www.univ-poitiers.fr
Institutions
Review
I have always been fascinated by knowledge about the evolution of man. Questions like ‘Where do we come from?’ and ‘Who was our common ancestor?’ are often posed by both pupils and scientists. This article describes a method that helps to strengthen the hypothesis of the hominoid evolution: that our forebears originated in Africa, after which they diversified in Asia, and repopulated Africa and Europe. This article could be used in biology lessons at the highest level, for pupils aged 16-18. It would be very useful when teaching general evolution and human evolution in particular. The article could also be used in physics class to show how advanced use of X-rays can be a valuable tool for biologists.
Sølve Tegnér Stenmark, Norway