




A grande migração Understand article
Traduzido por Artur Melo. Porque são as células parecidas com os gnus? Laura Spinney investiga a migração de células e a formação de órgãos, usando o pequeno e transparente peixe-zebra.
Todos os anos em Maio, enormes manadas de gnus se deslocam das planícies secas da África Oriental para as florestas, em busca de alimento. Regressam em Novembro, quando os aguaceiros vêm regar a planície, completando assim a sua migração anual. Cada gnu comporta-se de acordo com o seu instinto de sobrevivência, que tende a empurrá-lo para a companhia de outros gnus. Assim que se forma uma manada, ela comporta-se como uma nova unidade, com as suas próprias regras. As células têm um instinto de manada semelhante. São geneticamente programadas para interagir com outras células, mas não existe um conjunto específico de instruções que determine o comportamento preciso de cada célula no grupo. “Percebemos agora que muitos dos acontecimentos que ocorrem após a fecundação são de alguma forma livres e plásticos, e baseados em princípios de auto-organização,” diz Darren Gilmour do ‘European Molecular Biology Laboratory’ em Heidelberg, Alemanha.
Como biólogo do desenvolvimento, Darren gostaria de compreender esses princípios, porque a migração em massa de células dá origem à formação de sistemas intrincados e complexos, tais como os vasos sanguíneos e o sistema respiratório. Uma vez que as células que constituem estes grupos em migração parecem ser muito semelhantes, em termos dos genes que expressam, a supressão (desactivação) ou eliminação (redução da actividade de) de cada um dos genes tende a bloquear a migração de todo o grupo. Ele precisa de optar por uma abordagem mais subtil, descobrindo formas de bloquear as redes sociais das células e verificar como elas influenciam a sua capacidade de migrar e construir os órgãos. “É sociologia, basicamente,” diz ele. “Queremos saber quem diz a quem o que fazer, e como.”

Imagem cortesia de Darren Gilmour, EMBL
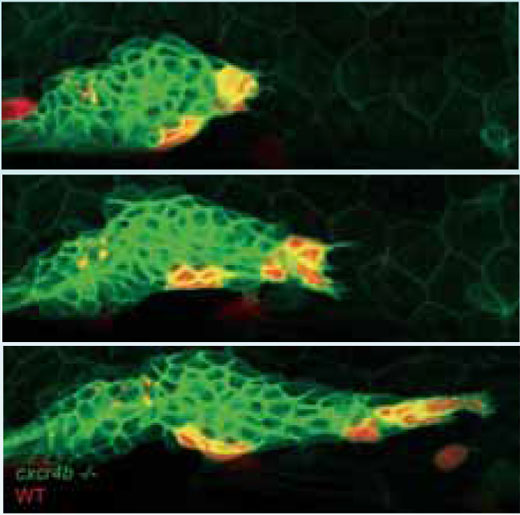
O grupo de Darren explora estes princípios no ‘primórdio’ da linha lateral do peixe-zebra, uma estrutura transitória que apenas pode ser observada no início do desenvolvimento, e cuja função é equipar o embrião do peixe com órgãos capilares – órgãos sensoriais que lhe permitem detectar a presença de outros peixes com base em pequenas variações na pressão da água. “O primórdio é um aglomerado de mais de 100 células que ‘rasteja’ sob a pele do animal, como uma lesma,” explica Darren. “Começa atrás do ouvido, dirigindo-se para a cauda, e à medida que avança deixa um rasto com cerca de duas células de espessura, marcado por pequenos grupos de células que se transformarão em órgãos capilares.”
À medida que os sistemas modelo evoluem, o peixe-zebra é uma resposta às preces dos biólogos, pois é transparente e consequentemente adequado para obtenção de imagens ao vivo de células marcadas com proteínas fluorescentes. O sistema da linha lateral é relativamente fácil de modificar geneticamente, e é também uma dádiva para quem se interessa por migração celular, porque utiliza uma via de sinalização que é muito importante na saúde humana, e, por isso, bem estudada. Um sinalizador químico chamado SDF1, o equivalente no peixe-zebra de um sinalizador também presente em humanos, provoca uma gama de alterações nas células do primórdio. Consegue-o ligando-se a um receptor, chamado Cxcr4b, que as células exibem no seu exterior. O Cxcr4b é uma proteína que acompanha a espessura da membrana celular e, em humanos, é necessária para ocorrer infecção pelo HIV. Além de desempenhar um papel no desenvolvimento normal, está também implicada na formação de metástases, ou propagação das células tumorais.
Num peixe-zebra mutante com reduzida quantidade de sinalizadores Cxcr4b, o grupo de Darren descobriu que o primórdio perde toda a orientação – não existe movimento de avanço e não se forma nenhum ‘rasto de lesma’. Além disso, a estrutura estática parece deixar de produzir grupos de células, como se tivesse que eliminar esses grupos para conseguir produzir mais. Um dos aspectos mais interessantes do mutante, no entanto, é que apesar da migração do primórdio ser desacelerada, uma observação mais atenta revela que as suas células ainda estão em movimento, chocando de maneira aleatória umas com as outras.w1 ”É como se estivessem confusas,” diz Darren. “Como gatos dentro de um saco.” E isso, acrescenta, é a chave para o funcionamento do sistema.
Para descobrir se podiam salvar o mutante e repor o movimento de avanço no primórdio, os investigadores criaram um mosaico genético transplantando algumas células normais, “com visão”, que expressaram o receptor Cxcr4b e podiam, por isso, reagir ao SDF1a, para o tecido mutante “cego”, com características opostas. As células normais e as mutantes foram marcadas com proteínas fluorescentes vermelhas e verdes, respectivamente, para que Darren e a sua equipa pudessem seguir o movimento das células. Rapidamente, usando o movimento às cambalhotas das células no primórdio, as células “com visão” encontraram o seu caminho para a frente, e o primórdio avançou novamente.
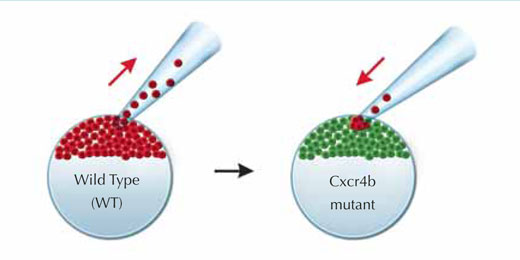
Imagem cortesia de Petra Riedinger, EMBL
do tipo ‘selvagem’ (vermelho)
encontram o seu caminho para a
extremidade do primórdio e
projectam-se na direcção da
migração, arrastando as células
mutantes (verde) com elas
Imagem cortesia de Darren
Gilmour, EMBL
O estranho nestas experiências, diz Darren, foi que nem todas as células sensoriais, com expressão do Cxcr4b, encontraram o seu caminho. Assim que algumas lá chegaram, o movimento desordenado terminou, o primórdio avançou, e aquelas células sensoriais que não conseguiram chegaram à frente permaneceram onde estavam – “como se estivessem congeladas”. “Pensamos que o sistema encontra força na instabilidade,” diz ele. “Anda às voltas em movimentos aleatórios, e, ao fazê-lo, asseguram que as células líderes se colocam em posição de poderem liderar. Sempre que elas puxam, as outras param.”
Mas se nem todas as células sensoriais se transformam em líderes, o que define uma líder? Seja o que for, não parece ser diferenças na actividade dos genes. ”Não são os genes que determinam, tu és a líder e tu és a que obedece,” diz Darren. Pelo contrário, os genes equipam todas as células com o receptor, e depois é uma questão de ‘luta entre elas’.” Ele e a sua colaboradora Petra Haas acham que a liderança é mais provavelmente definida pela natureza instável do grupo migratório. O movimento desordenado das células dá a algumas uma propensão temporária sobre as outras, em termos da sua proximidade à origem do sinal. Depois agarram-se a isso, e se forem capazes de manter a activação do seu receptor, avançam. Contudo, isso não é de modo nenhum o fim da história.
Uma teoria sobre migração de células considera que as células líderes levam atrás de si as que as seguem, as quais se deixam arrastar passivamente. Contudo, Darren é céptico sobre o que apelida de modelo ‘à boleia’. Ele prefere pensar que as líderes apontam às outras a direcção certa, e o primórdio é internamente organizado de tal forma que as outras as seguem activamente. E acredita que um princípio semelhante se aplica ao desenvolvimento de todos os órgãos. “Se olharmos para a ramificação dos vasos sanguíneos, prevejo que haverá um pequeno número de células na extremidade, que detectam os sinais do ambiente envolvente, e através de interacções célula-a-célula, influenciam de alguma forma o comportamento das outras,” diz ele. Apresenta a analogia de um falcão a emboscar um bando de estorninhos. Nem todos os estorninhos precisam de ver o falcão para reagir apropriadamente no voo – apenas precisam detectar o pânico dos seus vizinhos.
Até aqui, tudo bem, mas a história torna-se ainda mais complicada, pois verifica-se que nem todas as células seguidoras se comportam da mesma maneira. Nas suas experiências de imagens ao vivo, Darren e a sua equipa perceberam que à medida que a extremidade do primórdio abre caminho para avançar, as células na retaguarda abrandam, provocando o alongamento do tecido. O aluno de doutoramento Guillaume Valentin já identificou outro receptor, expresso por essas células da retaguarda, que também reage ao SDF1a, mas que desencadeia um tipo diferente de comportamento do suscitado pela via SDF1a/Cxcr4b – dando às células da retaguarda um certo grau de independência relativamente às células líder que expressam o Cxcr4b.
Quando o primórdio está a avançar, depositando grupos de células atrás de si, o movimento geral de células ao longo da estrutura é para trás. À medida que as células retrocedem, Darren suspeita que elas podem activar este segundo receptor. Talvez, especula ele, a combinação das duas populações de células que expressam receptores diferentes dêem origem a um movimento ‘de lagarta’, em que a frente e a retaguarda do primórdio avançam alternadamente. Algumas provas deste movimento provêm de experiências com embriões de peixe-zebra, nos quais a actividade do segundo receptor é reduzida e a falta a segunda metade desse movimento coordenado. “A extremidade que lidera puxa e puxa, mas a retaguarda não a acompanha, desiste e rola para trás,” diz ele.
Se estiver correcto, as novas descobertas levantam questões sobre o processo da transmissão de informação dinâmica através de grandes grupos de células. Torna-se necessário compreender não apenas a sinalização, mas também a propagação de forças através das populações de células, em termos de interacções célula-a-célula. É neste campo que Darren espera realizar a sua investigação. Por agora, no entanto, está entusiasmado pela descoberta de um segundo receptor SDF1a no primórdio do peixe-zebra. “A ideia de que as células internas também podem sentir é completamente original,” diz ele. Se o mesmo princípio se aplica aos gnus é outro assunto.
Resources
- Haas P, Gilmour D (2006) Chemokine signaling mediates self-organizing tissue migration in the zebrafish lateral line. Developmental Cell 10: 673-680. doi: 10.1016/j.devcel.2006.02.019
Institutions
Review
Fiquei encantada ao ser solicitada para comentar este artigo, pois poderia não o ter lido tão cuidadosamente de outra forma. Está muito bem escrito, com muitas analogias esclarecedoras, como a relação entre a migração de manadas de animais africanos e a migração de células em organismos em desenvolvimento. Além da biologia que é fascinante, incluindo proteínas sinalizadoras, infecção pelo HIV, peixes-zebra, células cancerosas e células mutantes ‘cegas’, também o é a descrição da metodologia da equipa de investigação, que se lê como uma história de detectives: apresentando teorias, rejeitando modelos, avançando para novas ideias e testando-as.
Há bastante interesse neste artigo para a maioria dos professores de biologia. Pode também constituir um exercício de compreensão realmente bom ou fornecer material suplementar para alunos de biologia com idades a partir dos 16 anos. Em alternativa, pode ser usado num debate sobre embriologia, desenvolvimento de vertebrados, engenharia genética, comunicação entre células em oposição a comunicação entre organismos, proteínas sinalizadoras, cancro, mutações, as fases das investigações científicas ou modelação de teorias.
Sue Howarth, Reino Unido