Dobór naturalny molekularnie Understand article
Tłumaczenie Jadwiga Schreiber. Wiadomo, że niektóre sekwencje genetyczne mogą pomóc organizmom przetrwać w ich środowisku. Jednak wykazanie, które sekwencje genetyczne są korzystne i w jaki sposób pomagają nam przeżyć nie jest łatwe - zwłaszcza w naturalnych populacjach zwierząt…

iStockphoto
Kiedy ludzie po raz pierwszy opuścili Afrykę około 150 000 lat temu i zaczeli osiedlać się w dolinach Tygrysu i Eufratu, żeglować między wyspami Indonezji czy wędrować przez Cieśninę Beringa do Ameryki, napotkali wiele wyzwań. Populacje pochodzące z gorącej, suchej sawanny afrykańskiej musiały przystosować sie do lokalnych warunków środowiska. W ciągu wielu pokoleń ich fizjologia i wygląd uległy przystosowawczym zmianom (Harris & Meyer, 2008): skóra ludzi stała się jaśniejsza gdy zamieszkali oni w mniej nasłonecznionych regionach (Lamason et al., 2005); populacje, które piły mleko zwierząt domowych zachowały zdolność do trawienia laktozy również w dorosłym wieku (cechę traconą krótko po wieku niemowlęcym w populacjach nie znających mleka zwierząt hodowlanych (Tishkoff et al., 2007); populacje, które spożywały produkty bogate w skrobię produkowały więcej amylazy w ślinie, enzymu który pomaga w trawieniu skrobi (al Perry et al., 2007).
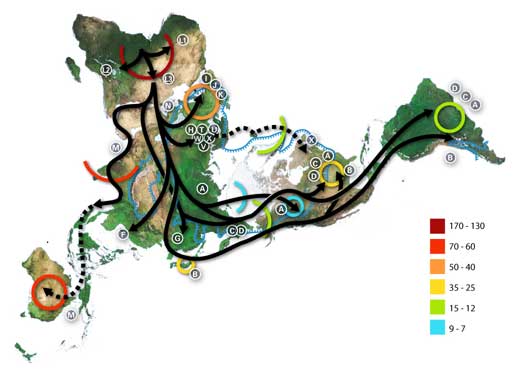
Niebieska linia reprezentuje obszar porkyty lodem lub tundrą w czasie ostatniej epoki lodowcowej.
Litery oznaczają tzw. haplogrupy mitochondrialnego DNA (mtDNA), które wykorzystywane są do genetycznej i geograficznej charakteryzacji populacji ludzkich.
Na przykład, typowy podział haplogroup mtDNA jest następujący:
Afrykański: L, L1, L2, L3, L3
Bliskowschodni: J, N
Południowo europejski: J, K
Ogółno europejski: H, V
Północno europejski: T, U, X
Azjatycki: A, B, C, D, E, F, G (M składa sie z C, D, E i G)
Północnoamerykański: A, B, C, D, i czasem X.
Click to enlarge image
Zdjęcia dzięki uprzejmości Avsa; źródło: Wikimedia Commons
Uważa się, że niektóre z tych zmian są konsekwencją pozytywnego doboru naturalnego (zobacz słowniczek dla wszytkich słów napisanych kursywą). Oznacza to, że w przeszłości w specificznym środowisku pojedyncze osobniki przypadkowo posiadające korzystną sekwencję DNA przeżyły i zostawiły po sobie wiecej potomstwa niż inne osobniki, które posiadały inną, mniej korzystną sekwencję DNA. Dziś, wykorzystując sekwencje genomowe wielu gatunków, w tym ludzi i ich najbliższych ewolucyjnych krewnych, naukowcy mogą porównać sekwencje DNA i cechy organizmów pochodziących z grup (populacji lub gatunków) posiadających różne style życia i żyjących w różnych środowiskach aby zidentyfikować sekwencje, które mogły odgrywać rolę w adaptacjach. To z kolei pozwala im badać funkcje DNA i jego potencjału adaptacyjnego dla organizmu.

JBryson / iStockphoto
Niektóre geny wpływające na kolor skóry u ludzi wykazują geograficznie specyficzną zmianę w sekwencji; porównania pomiędzy populacjami europejskimi i afrykańskimi sugerują, że różnorodność koloru skóry jest skutkiem pozytywnego doboru naturalnego. Jasność skóry koreluje ze wzrostem szerokości geograficznej.
Jedna z hipotez wyjaśniających to zjawisko głosi, że jasna skóra sprzyja produkcji witaminy D i poparta jest obserwacjami, że ludzie o ciemnej skórze żyjący na wysokich szerokościach geograficznych cierpią na niedobór witaminy D. Ponadto, jasna skóra jest bardziej wrażliwa na szkodliwe działanie światła słonecznego: większa ekspozycja koreluje ze zwiększonym ryzykiem wystąpienia raka skóry u ludzi o jasnej karnacji. Dlatego też jasny kolor skóry u ludzi żyjących na wysokich szerokoścach geograficznych może być ewolucyjnym kompromisem pomiędzy ochroną przed rakotwórczym działamien promieni słonecznych i możliwością produkcji niezbędnej witaminy.
mikroskopu skaningowego
przedstawiające krwinki
sierpowate i zdrowe krwinki
czerowne
Zdjęcia dzięki uprzejmości EM
Unit, UCL Medical School, Royal
Free Campus / Wellcome
Images
Choć hipoteza ta jest przekonująca, dowód jej słuszności jest pośredni. Bezpośrednie wykazanie adaptacyjnej wartości tej cechy wymagałoby potwierdzenia, że na wyższych szerokościach geograficznych osobniki z jasną skórą mają większe szanse na przeżycie i reprodukcję. Takie stwierdzenie w przypadku naszego gatunku jest jednak trudne: przetestowanie zdolności przeżycia (test, w którym osobiki o różnych cechach są skonfrontowane z pewnym środowiskiem w celu zbadania który z nich przetrwa lepiej/dłużej) nie jest możliwe, a długi czas naszego życia utrudnia zbadanie różnic reprodukcyjnych. Okoliczności, w jakich możemy zaobserwować cechę ukształtowaną dzięki doborowi naturalnemu u ludzi są zatem bardzo ograniczone – ale istnieją.
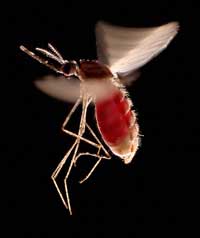
krwi. Gatunek ten, Anopheles
stephensi, jest nosicielem
malarii w Indiach i Pakistanie
Zdjęcia dzięki uprzejmości
Hugh Sturrock / Wellcome
Images
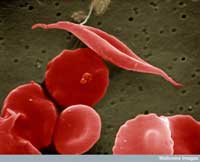
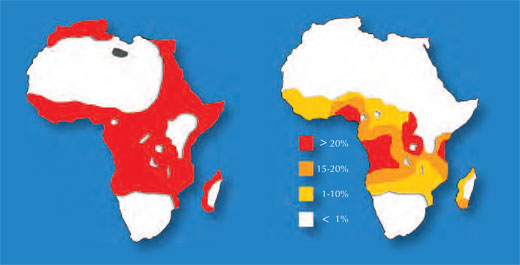
Jeden z przykładów dotyczy dwóch chorób: anemii sierpowatej i malarii. Gen zaangażowany w anemię sierpowatą występuje w dwóch odmianach, tzw. allelach: „normalny” allel i allel anemii sierpowatej. Osobniki z dwoma allelami anemii sierpowatej cierpią na ciężką amemię, podczas gdy osobniki z jednym allelem normalnym i jednym allelem anemii sierpowatej nie wykazują poważnych objaw tej choroby. Statystyki zgonów sugerują, że posiadanie allelu anemii sierpowatej może być korzystne: w populacjach narażonych na malarię osobniki posiadające jeden allel anemii sierpowatej i jeden allel normalny mają większą szansę przeżycia niż osobniki z dwoma normalnymi allelami, ponieważ pasożyt wywołujący malarię (zarodziec sierpowaty (Plasmodium falciparum)) wymaga zdrowych komórek krwi by je zasiedlić i się rozmnożyć. Dlatego też częstość występowania allelu anemi sierpowatej wzrasta u ludzi narażonych na zakażenie malarią – allel ten umożliwia adaptację w tym środowisku.
Zdjęcia dzięki uprzejmości Anthony Allison; źródło: Wikimedia Commons
Innym przykładem demonstrującym adaptacyjne źródło cechy człowieka jest fragment chromosomu 17, który uległ odwróceniu u naszych przodków ponad trzy miliony lat temu (Stefansson et al., 2005). Fakt, że ten odwrócony fragment chromosomu 17 rozprzestrzenił się w populacjach Europy sugeruje, że musiał on mieć (pośrednio lub bezpośrednio) korzystny wpływ na osobniki, które go posiadały. Ustalając genotyp prawie 30 000 Islandczyków, naukowcy byli w stanie wykazać, że przez ostatnie 80 lat osoby posiadające odwrócony fragment chromosomu 17 pozostawiły po sobie średnio 3,2% potomstwa na pokolenie więcej niż osobinki z „normalną” sekwencją, tłumacząc tym samym szybkie rozprzestrzenienie się odwróconej wersji.
Choć te dwa przyklady wyraźnie wskazują na istnienie pozytywnej selekcji u ludzi, molekularne mechanizmy wpływu zmienności sekwencji genetyczych na fenotyp u ludzi nie są jasne i wymagają rozpatrywania każdego przypadku z osobna. Aby wyjaśnić związki przyczynowo-skutkowe pomiędzy potencjalnie korzystną sekwencją DNA i dostosowaniem (ang. fitness) osobnika, naukowcy kierują się w stronę organizmów nieco łatwiejszych w przeprowadzaniu eksperymentów niż człowiek.
Kolor sierści myszy Peromyscus polionotus podobny jest do koloru podłoża, na którym żyją, zapewniając kamuflaż. Myszy żyjące na jasnym piasku plaż na Florydzie mają dużo jaśniejszą sierść niż myszy tego samego gatunku, żyjące w środkowej części kraju na brązowo-szarej ziemi. Adaptacyjną wartość tej cechy udowodniono ponad 30 lat temu: myszy z sierścią podobną do podłoża były dużo rzadziej zjadane przez sowy niż myszy, których kolor sierści nie odpowiadał podłożu. Mimo to dopiero nieadawno naukowcy odkryli sekwencje odpowiadające za tę cechę (Hoekstra et al., 2006): odmiany koloru sierści w dużej mierze zależą od rożnych allelów genu McR1. Białko kodowane przez ten gen funkcjonuje jak biochemiczny przełącznik pomiędzy produkcją eumelaniny, ciemnego barwnika skóry i feomelaniny, jasnego barwnika skóry. Różne allele genu McR1 aktywują w różnym stopniu proces produkcji pigmentu skóry, zwiększając produkcję jednego pigmentu bardziej niż innego.
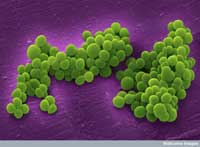
mikroskopu skaningowego
przedstawiające skupisko
gronkowców złocistych
(Staphylococcus aureus)
opornych na metycilinę
Zdjęcia dzięki uprzejmości
Annie Cavanagh / Wellcome
Images
Innym przykładem jest oporność gronkowca złocistego (Staphylococcus aureus) na leki, bakterii wywołującej m.in. zapalenie płuc lub zapalenie zastawek serca. W rzadko występujących naturalnych eksperymentach, pacjentowi z powracjącymi zakażeniami gronkowcem złocistym (S. aureus) podawano przez trzy miesiące wankomycynę, jeden z niewielu antybiotyków, które są jeszcze skuteczne w zwalczaniu tej bakterii. Przed i w trakcie leczenia naukowcy izolowali próbki patogenu i zsekwencjonowali cały genom pierwszej i ostatniej próbki. Kiedy porównali oni trzy miliony par zasad („liter” kodu genetycznego) z jakich zbudowne jest DNA tej bakterii, znaleźli oni tylko 35 różnic pomiędzy pierwszą i ostatnią wyizolowaną bakterią.
Częściowo sekwencjonując genom izolatów pośrednich naukowcy odtworzyli kolejność w jakiej te zmiany nastąpiły. Testując in vitro oporność różnych izolatów tej bakterii na wankomycynę, skorelowali poszczególne zmiany genetyczne wpływające na wzrost bakterii i jej reakcji na lek. Na przykład, pierwszy i drugi szczep tej bakterii rożnił się sześcioma nukleotydami (zmianami w „literach” DNA) w dwóch genach. Mutacje te były wyjątkowo korzystne: zwiększyły one tolerancję tej bakterii na wankomycynę czterokrotnie, pozwalając bakterii posiadającej te mutacje przeżyc i lepiej się rozmnożyć, rozprzestrzeniając w ciele chorego. Dwadzieścia sześć kolejnych mutacji, jakie wystąpiły w kolejnych tygodniach leczenia, jeszcze podwoiły oporność, prowadząc do powstania gronkowców tolerujących wankomycynę (Mwangi et al., 2007).
Podsumowując, badanie molekularnego podłoża ewolucji adaptacyjnej u dzikich populacjach nie jest łatwe. Wymaga zidentyfikowania presjii selekcyjnych działających na organizmy w danym środowisku, zmierzenia dostosowania osobników, rozpoznania sekwencji DNA (jednej lub wielu) odpowiadającej za daną cechę i wyjaśnienia mechanizmu w jaki sposób zmiany w tej sekwencji wpływają na dostosowanie. Jednakże wykorzystując organizmy modelowe i najnowsze osiągnięcia techniczne badania te stają sie coraz bardziej dostępne, zwiększając naszą wiedzę dotyczącą molekularnych podstaw adaptacji organizmów w środowisku naturalnym.
Słowniczek
Adaptacja: dana cecha jest adaptacją jeśli umożliwia ona osobnikowi (który ją posiada) lepiej przetrwać (przeżyć) i reprodukować się w danym środowisku w porównaniu z osobnikami, które tej cechy nie posiadają. Bardziej formalnie, dana cecha jest adaptacją jeśli zwiększa dostosowanie (z ang. fitness) osobnika.
Allel: wariant genu.
Dostosowanie (z ang. fitness): trudny do zdefiniowania termin z dziedziny biologii ewolucyjnej i genetyki populacji; określa on średnią liczbę potomstwa w ciągu jednego pokolenia przypadającą na konkretny genotyp w porównaniu z innym genotypem w populacji. Genotypy, które pozostawiają więcej potomstwa są lepiej dostosowane. Dobry przęgląd informacji na temat dostosowania i genotypów można znaleźć w Wikipediiw1.
Genom: całkowite DNA danego organizmu. Zazwyczaj odnosi się to do jądrowego DNA u organizmów eukariotycznych, ale obejmuje też DNA mitochondrialne czy plastydowe, lub całość materiału genetycznego organizmów prokariotycznych i wirusów. Więcej informacji na ten temat można znaleźć w „What is a genome” na stronie internetowej Amerykańskiej Nacjonalnej Biblioteki (US National Library)w2.
Dobór naturalny: jest jednym z mechanizmów ewolucji, opisuje ona różnice w przetrwaniu i reprodukcji osobników w danym środowisku. Dobór naturalny jest ‘pozytywny’, kiedy promuje cechę pozwalając osobnikom, które ją posiadają, lepiej przetrwać i rozmnażać się niż osobnikom nie posiadającym tej cechy.
Presja selekcyjna: czynnik środowiskowy (np. temperatura, występowanie pasożytów, drapieżnictwo lub agresja ze strony osobników tego samego gatunku) prowadzący do zróżnicowania szansy przeżycia i reprodukcji organizmów.
Cecha: jedna lub zespół właściwości danego organizmu (np. wzrost, odporność na antybiotyki, zdolność rozpoznawania kolorów czy umiejętność zwijania języka w rurkę).
Podziękowania
Autor składa podziękowania Davidowi Hughesowi, Mehmetowi Somelowi i Ani Lorenc za pomocne komentarze do tego artykułu.
References
- Harris EE, Meyer D (2006) The molecular signature of selection underlying human adaptations. American Journal of Physical Anthropology 131(S43): 89-130. doi: 10.1002/ajpa.20518
- Artykuł ten stanowi dobry przegląd na temat badań ewolucji molekularnej u ludzi.
- Hoekstra H et al. (2006) A single amino acid mutation contributes to adaptive beach mouse color pattern. Science 313: 101-104. doi: 10.1126/science.1126121
- Ten i inne artykuły dotyczące umaszczenia myszy opublikowane przez grupę naukową Hopi Hoekstra dostępne są na stronie internetowej Uniwersytetu Harwarda. Zobacz: www.oeb.harvard.edu/faculty/hoekstra/Links/PublicationsPage.html
- Zobacz również uzupełniający artykuł, w którym opisane jest odkrycie jak gen Agouti, negatywny regulatora genu McR1, przyczynia się do powstania maskującej barwy sierści u myszy Peromyscus:
- Steiner CC, Weber JN, Hoekstra HE (2007) Adaptive variation in beach mice produced by two interacting pigmentation genes. PLoS Biology 5: e219. doi: 0.1371/journal.pbio.0050219
- Ten i wszystkie inne artykuły z PLoS Biology dostępne są bezpłatnie online.
- Podany niżej artykuł opisuje adaptacyjną pigmentację u kręgowców:
- Hoekstra HE (2006) Genetics, development and evolution of adaptive pigmentation in vertebrates. Heredity 97: 222-234. doi: 10.1038/sj.hdy.6800861
- Artykuł ten można bezpłatnie pobrać z strony internetowej czasopisma Heredity: www.nature.com/hdy
- Przegląd badań Hopi Hoekstra wymienionych w artykule dostępny jest na blogu Johna Hawksa: http://johnhawks.net/weblog/topics/evolution/selection/hoekstra-2009-adaptive-pigmentation.html
- Lamason RL et al. (2005) SLC24A5, a putative cation exchanger, affects pigmentation in zebrafish and humans. Science 310: 1782-1786. doi: 10.1126/science.1116238
- Mwangi MM et al. (2007) Tracking the in vivo evolution of multidrug resistance in Staphylococcus aureus by whole-genome sequencing. Proceedings of the National Academy of Sciences of the United States of America 104: 9451-9456. doi: 10.1073/pnas.0609839104
- Perry GH et al. (2007) Diet and the evolution of human amylase gene copy number variation. Nature Genetics 39: 1256-1260. doi: 10.1038/ng2123
- Zobacz również przegląd tych badań w Panda’s Thumb: http://pandasthumb.org/archives/2008/12/amylase-and-hum.html
- Stefansson H et al. (2005) A common inversion under selection in Europeans. Nature Genetics 37: 129-137. doi: 10.1038/ng1508
- Zobacz także omówienie nowszych danych dotyczące tego regionu, sugerujące bardziej skomplikowane wyjaśnienie jego rozprzestrzenienia się: http://evolgen.blogspot.com/2005/02/human-inversion-under-selection.html
- Tishkoff SA et al. (2006) Convergent adaptation of human lactase persistence in Africa and Europe. Nature Genetics 39: 31-40. doi: 10.1038/ng1946
- Zobacz również przegląd tych badań w The New York Times: www.nytimes.com/2006/12/10/science/10cnd-evolve.html?_r=1
Web References
- w1 – dobry opis pojęć takich jak dostosowanie (‘fitness’) czy ‘genotyp’ można znaleźć w Wikipedii: http://en.wikipedia.org/wiki/Fitness_(biology) i http://en.wikipedia.org/wiki/Genotype
- w2 – Więcej informacji na temat genomu i Projektu ‘Human Genome Project’ jest dostępnych w ‘What is a genome’ na stronie internetowej Amerykańskiej Narodowej Biblioteki Medycznej (US National Library of Medicine): http://ghr.nlm.nih.gov/handbook/hgp/genome
Resources
- Jeśli ten artykuł Ci się spodobał, może zainteresują Cię również inne artykuły o ewolucji w „Science in School”:
- Haubold B (2010) Review of Why Evolution is True. Science in School 14. www.scienceinschool.org/2010/issue14/evotrue
- Leigh V (2008). Interview with Steve Jones: the threat of creationism. Science in School 9: 9-17. www.scienceinschool.org/2008/issue9/stevejones
- Patterson L (2010) Getting ahead in evolution. Science in School 14: 16-20. www.scienceinschool.org/2010/issue14/amphioxus
- Pongsophon P, Roadrangka V and Campbell A (2007) Counting Buttons: demonstrating the Hardy-Weinberg principle. Science in School 6: 30-35. www.scienceinschool.org/2007/issue6/hardyweinberg
- Więcej informacji na temat malarii można znaleźć:
- Hodge R (2006) Fighting malaria on a new front. Science in School 1: 72-75. www.scienceinschool.org/2006/issue1/malaria
- Aby dowiedzieć się więcej na temat struktury skrobi rozkładanej min. przez ślinowe amylazy zobacz:
- Cornuéjols D (2010) Starch: a structural mystery. Science in School 14: 22-27. www.scienceinschool.org/2010/issue14/starch
Review
Artykuł ten opisuje szereg interesujących przykładów ewolucyjnych adaptacji na poziomie molekularnym u ludzi. Podkreśla on również trudności w wyjaśnieniu związków pomiędzy adaptacyjnym DNA i dostosowaniem (‘fitness’) ludzi oraz potrzebę zastosowania innych organizmów modelowych w tych badaniach.
Artykuł ten doskonale nadaje się jako materiał sprawdzający rozumienie tematu i pytań dotyczących doboru naturalnego i dostodowania (‘fitness’) ludzi i zwierząt modelowych. Na przykład:
- Wyjaśnij procesy zaangażowane w dobór naturalny (selekcję naturalną).
- Co rozumiesz pod pojęciem ‘dostosowanie’ (z ang. ‘fitness’)?
- Wyjaśnij w jaki sposób allel anemii sierpowatej może być korzystny u niektrórych populacji ludzkich.
- Jakie problemy związane są ustaleniem związków przyczynowo skutkowych pomiędzy adaptacyjnym DNA i dostosowaniem u ludzi?
- Narysuj schemat wyjaśniający adaptacyjną wartość koloru sierści myszy Peromyscus polionotus.
- W jaki sposób naukowcy byli w stanie powiązać zmiany genetyczne w gronkowcu złocistym (Staphylococcus aureus) z jego wzrostem i odpornością na antybiotyki?
Artykuł ten umożliwia również studentom zbadanie powiązań pomiędzy DNA, sekwencją aminokwasów, strukturą białka i jego funkcją w anemii sierpowatej. Tekst ten nadaje się do lekcyjnej dyskusji na temat metod i problemów związanych z badaniami molekularnych podstaw zależności ewolucyjnych i etyki badań genetycznych w populacji ludzkiej. Interdyscyplinarne badania mogłyby być przeprowadzone łącząc takie dziedziny jak np. historię nauki i genetykę populacji.
Mary Brenan, Wielka Brytania