Natürliche Selektion auf molekularer Ebene Understand article
Übersetzt von: Veronika Ebert, Höhere Bundeslehr- und versuchsanstalt für chemische Industrie, Wien Wir wissen, dass bestimmte genetische Sequenzen für das Überleben in unserer Umwelt nützlich sein können – das ist die Basis der Evolution. Es ist jedoch alles andere als leicht heraus zu…

iStockphoto
Als die Menschen zu Beginn, vor etwa 150 000 Jahren, Afrika verließen, sich in den Tälern von Euphrat und Tigris ansiedelten, zwischen den indonesischen Inseln segelten, die Beringstraße überquerten, um schließlich Amerika zu erreichen, hatten sie viele Schwierigkeiten zu bestehen. Aus den heißen, trockenen Savannen Afrikas stammend, mussten sie sich an lokale Umweltbedingungen anpassen. Ihre Physiologie und ihr Aussehen änderte sich über viele Generationen demensprechend (Harris & Meyer, 2008). Die Haut jener Menschen, die in sonnenärmeren Gegenden lebten, wurde heller (Lamason et al., 2005). Populationen, die Milch von Haustieren konsumierten, behielten die Fähigkeit Milchzucker (Laktose) abzubauen bis in das Erwachsenenalter bei. Diese Fähigkeit geht nach der Kindheit verloren, wenn keine Milch getrunken wird (Tishkoff et al., 2007). Populationen, die Stärke-reiche Lebensmittel verzehrten, erzeugten höhere Mengen von Speichelamylase, dem Enzym, das an der Verdauung von Stärke beteiligt ist (Perry et al., 2007).
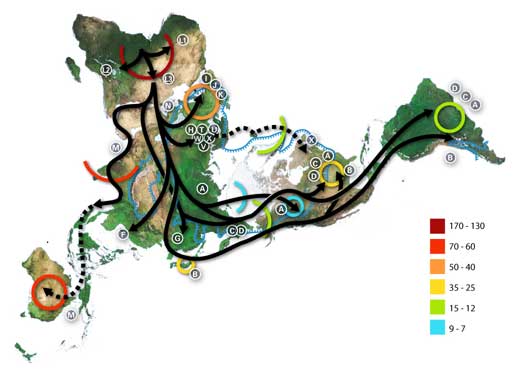
Die Zahlen geben die Zeitspanne als Vielfaches von Tausend vor der Jetztzeit an.
Die blauen Linien begrenzen die Areale, die während der letzten Eiszeit entweder von Eis bedeckt, oder Tundra waren. Die Buchstaben stehen für DNA-Haplogruppen (reine mütterliche Linien). Haplogruppen können als genetische Populationen definiert werden, sie zeigen oft eine regionale Verteilung.
Beispiele für die regionale Verteilung bestimmter Haplogruppen mitochondrialer DNA:
Afrika: L, L1, L2, L3, L3
Naher Osten: J, N
Südeuropa: J, K
Europa gesamt: H, V
Nordeuropa: T, U, X
Asien: A, B, C, D, E, F, G (Anmerkung: M besteht aus C, D, E, and G)
Ureinwohner Amerikas: A, B, C, D, , gelegentlich auch X.
Zum Vergrößern auf das Bild klicken
Mit freundlicher Genehmigung von Avsa; Bildquelle: Wikimedia Commons
Zumindest ein Teil dieser Veränderungen dürfte auf eine positive Selektion (Erklärungen für kursiv geschriebene Begriffe siehe Glossar)zurückzuführen sein. Das bedeutet, dass bestimmte Umweltbedingungen (der Selektionsdruck) in der Vergangenheit dazu geführt hat, dass Menschen mit vorteilhaften Gensequenzen überlebt und eine größere Zahl von Nachkommen hervorgebracht haben als Menschen mit anderen, weniger vorteilhaften Ausprägungen. Die Untersuchung der Genomsequenz vieler Arten, darunter auch jene des Menschen und seiner nächsten Verwandten, mit modernen Untersuchungsmethoden erlaubt es, die Merkmale und DNA-Sequenzen von Populationen oder Arten mit unterschiedlicher Lebensweise und unter verschiedenen Umweltbedingungen zu vergleichen. Dadurch kann festgestellt werden, welche Sequenzen für die Anpassungen verantwortlich sein könnten. Die Forscher/innen können somit die Funktion der DNA-Abschnitte und ihre “adaptive Bedeutung” für ein Lebewesen ermitteln.

von JBryson / iStockphoto
Verschiedene Sequenzvarianten einiger Gene, die die Hautfarbe beim Menschen regeln, zeigen eine spezifische geographische Verteilung. Sequenzvergleiche zwischen europäischen und afrikanischen Populationen lassen vermuten, dass die Variabilität der Hautfarbe auf eine positive Selektion zurück zu führen ist. Helle Hauttönungen korrelieren positiv mit größerer geographischer Breite. Für diesen Effekt gibt es verschiedene Hypothesen, die den potentiellen Vorteil hellerer Haut erklären können.
Die Annahme, dass die Bildung von Vitamin D durch eine helle Hautfarbe gefördert wird, wird durch Beobachtungen an Menschen mit dunkler Hautfarbe unterstützt. Wenn letztere in größeren geographischen Breiten leben, leiden sie unter Vitamin D Mangel. Außerdem ist eine hellere Haut empfindlicher gegenüber der schädlichen Wirkung des Sonnenlichts: Stärkere Sonneneinstrahlung korreliert mit dem Auftreten von Hautkrebs bei hellhäutigen Menschen. Es ist daher anzunehmen, dass die helle Haut der in größeren geographischen Breiten lebendenden Menschen, einen evolutionärer Kompromiss zwischen dem Schutz vor den krebserregenden Effekten des Sonnenlichts und der ausreichenden Produktion des notwendigen Vitamin D´s darstellen.
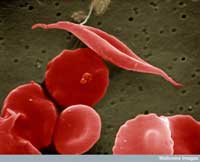
Aufnahme von Sichelzellen
und anderen, normalen roten
Blutzellen
Mit freundlicher Genehmigung
der EM Unit, UCL Medical
School, Royal Free Campus /
Wellcome Images
Obwohl es sich dabei um eine stichhaltige Hypothese handelt, ist sie dennoch nur indirekt belegbar. Der direkte Nachweis für den adaptativen Wert der Hautfarbe würde eine Messung erfordern, mit der festgestellt werden kann, ob hellhäutige Menschen in größerer Höhe länger überlegen und mehr Nachwuchs in die Welt setzen. Diese Nachweise sind in menschlichen Populationen schwer zu erbringen: Überlebensexperimente, bei denen Einzelne verschiedenen Umweltbedingungen ausgesetzt werden, um festzustellen, wer besser überleben kann, können mit Menschen nicht durchgeführt werden. Unsere lange Generationszeit macht es auch schwierig, unterschiedliche Reproduktionsraten zu detektieren. Obwohl die Bedingungen, unter denen es möglich ist, den adaptiven Wert eines Merkmals im Menschen zu erfassen begrenzt sind, gibt es sie doch.
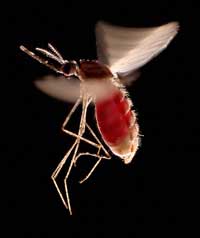
blutgefüllten Bauch. Diese
Art, Anopheles stephensi, ist
der Insektenwirt, der die
Malaria in Indien und
Pakistan überträgt
Bild zur Verfügung gestellt von
Hugh Sturrock / Wellcome
Images
Ein Beispiel umfasst zwei Krankheiten: die Sichelzellanämie und die Malaria. Das Gen, das bei der Sichelzellanämie betroffen ist, kommt in zwei Varianten, sogenannten Allelen vor: ein “normales” Allel und ein Sichelzellallel. Personen mit dem Sichelzellallel leiden unter einer schweren Sichelzellanämie, während jene mit dem normalen Allel gesund sind. Die Analyse von Mortalitäts-(Todes-)raten last vermuten, dass das Sichelzellallel trotzdem vorteilhaft sein kann: in Populationen, die mit dem Malariaerreger in Kontakt kommen, haben Personen, die über je ein normales Allel und ein Sichelzellallel verfügen, größere Überlebenschancen als Personen mit zwei normalen Allelen. Verantwortlich dafür dürfte sein, dass der Parasit (Plasmodium falciparum) intakte rote Blutzellen benötigt, in die er eindringen und sich vermehren kann. Daher nimmt die Häufigkeit des Sichelzellallels in Gruppen, die dem Malariaerreger ausgesetzt sind, zu – die Allelausstattung wird der Umwelt angepasst.
Mit freundlicher Genehmigung von Anthony Allison;
: Wikimedia Commons
Ein anderes Beispiel für den adaptiven Wert eines menschlichen Merkmals betrifft ein Fragment des Chromosoms 17, das in unseren Vorfahren vor mehr als drei Millionen Jahren invertiert (umgedreht) worden ist (Stefansson et al., 2005). Die Tatsache, dass sich diese Variante in den europäischen Populationen verbreitet hat, lässt vermuten, dass hier eine positive Selektion stattfand –Individuen mit dieser Variante hatten Vorteile. Die Genotypisierung der meisten Isländer/innen zeigte, dass Individuen mit dieser Sequenzvariante während der letzten 80 Jahre durchschnittlich 3,2% mehr Nachkommen pro Generation hatten als Personen mit der normalen Sequenz. Eine plausible Erklärung, wie sich diese Variante so rasch verbreiten konnte.
Obwohl diese Beispiele die Wirkung erst kürzlich stattgefundener positiver Selektion im Menschen klar belegen können, bleibt der molekulare Mechanismus, der erklärt, warum diese Sequenzvariationen Vorteile bringen können, im Dunklen. Hier muss jeder Einzelfall gesondert untersucht werden. Zur Aufklärung der kausalen Zusammenhänge zwischen mutmaßlichen adaptiven DNA-Abschnitten und der Fitness eines Individuums, haben sich die Wissenschafter/innen einfacher untersuchbaren Modellorganismen zugewandt.
Zum Beispiel der Alabama-Küstenmaus, Peromyscus polionotus. Die Farbe ihres Fells passt zur Farbe des jeweiligen Untergrunds und bietet dadurch eine exzellenteTarnung. Mäuse, die auf dem hellen Sand Floridas leben, sind viel heller als Individuen der gleichen Art, die im Landesinneren beheimatet sind. Der adaptive Wert dieses Merkmal konnte schon vor 30 Jahren experimentell gezeigt werden: Mäuse mit einer zum jeweiligen Untergrund passenden Fellfarbe wurden seltener von Eulen gefressen als schlechter getarnte Mäuse. Dennoch konnten erst kürzlich die für diese adaptive Merkmal verantwortlichen Genorte identifiziert werden (Hoekstra et al., 2006): Die Variation der Fellfarbe hängt in erster Linie von den unterschiedlichen Allelen des McR1 Gens ab. Das Protein, das von diesem Gen kodiert wird, fungiert als biochemischer Schalter, der entweder die Produktion von Eumelanin, dem dunklen Hautpigment, oder von Phaeomelanin, einem hellen Pigment, steuert. Die beiden Allele des McR1-Gens aktivieren die Pigment-produzierenden Stoffwechselwege in einem unterschiedlichen Ausmaß, wodurch entweder die Produktion des einen, oder des anderen Pigments begünstigt wird.
Aufnahme von Gruppen
Methicillin-resistenter
Staphylococcus aureus-
Bakterien
Mit freundlicher Genehmigung
von Annie Cavanagh /
Wellcome Images
Ein weiteres Beispiel für einen belegbaren kausalen Zusammenhangs bietet Staphylococcus aureus, ein Bakterium, das für Erkrankungen wie Lungenentzündung, oder Herzklappenentzündungen, verantwortlich ist. In einem selten vorkomenden natürlichen Experiment wurde ein Patient mit immer wiederkehrenden S. aureus-Infektionen drei Monate lang mit Vancomycin behandelt. Vancomycin ist eines jener wenigen Antibiotika, die bei S. aureus-Infektionen noch immer wirksam sind. Vor, und während der Behandlung wurden Proben (Isolate) des Krankheitserregers gesammelt. Die Isolate vom Erkrankungsbeginn und jene vom Ende der Infektion wurden komplett sequenziert. Der Vergleich der drei Millionen Basenpaare, der gesamten genetischen Information dieses Bakteriums, zeigte, dass nur 35 Basen verändert waren.
Durch Teilsequenzierung von Isolaten aus der mittleren Erkrankungsphase konnte die Reihenfolge, in der diese Basenveränderungen aufgetreten sein mussten, geklärt werden. Die in vitro Testung der Empfindlichkeit der verschiedenen Isolate gegenüber Vancomycin erlaubte es, die einzelnen Basensubstitutionen (Austausch einzelner Basen der DNA) und ihre Auswirkung auf die Resistenz des Erregers gegenüber dem Medikament zu korrelieren. So fand man zwischen dem ersten und dem zweiten Isolat sechs Unterschiede in der Nukleotidsubstitutionen. Diese sechs Mutationen waren eindeutig vorteilhaft: sie verstärken die Vancomycinresistenz des Bakteriums um den Faktor 4, und erhöhten dadurch die Überlebenswahrscheinlichkeit und Vermehrungsrate von Bakterien mit dieser Mutation. Der Anteil an mutierten Bakterien an der Gesamtpopulation hatte sich dadurch erhöht. Sechsundzwanzig weitere Mutationen während der nachfolgenden Behandlungswochen verdoppelten die Resistenz, sodass ein Vancomydin-resistenter Stamm von S. aureus enstehen konnte (Mwangi et al., 2007).
Zusammenfassend kann gesagt werden, dass es nicht einfach ist, die molekularen Ursachen für die adaptive Evolution in natürlichen Populationen aufzuklären. Es ist notwendig, den selektiven Druck zu definieren, die für das jeweilige Merkmal verantwortlichen DNA-Sequenzen zu identifizieren, und die Mechanismen aufzuklären, wie das adaptive Merkmal durch die jeweilige Sequenzveränderung beeinflusst wird. Erst durch die Verwendung von Modellorganismen und neuerer Untersuchungsmethoden sind solche Untersuchungen möglich geworden, wodurch unser Verständnis, wie bestimmte genetische Veränderungen die Anpassung von Lebewesen an ihre Umwelt ermöglichen, zugenommen hat.
Glossar
Adaptiver Wert: : eine Merkmal hat einen „adaptiven Wert“, wenn sie einem Individuum die Fähigkeit verleiht, unter bestimmten Umweltbedingungen zu überleben und sich stärker fortzupflanzen als Individuen ohne dieser Merkmal. Eine Merkmal ist „adaptiv“, wenn es die Fitness verbessert.
Allel: eine Genvariante
Fitness: ein schwer definierbarer Ausdruck aus der Evolutionsbiologie und Populationsgenetik. Er gibt die mittlere Zahl von Nachkommen über eine Generation an, die mit einem bestimmten Genotyp assoziiert ist und vergleicht diesen Wert mit jenem eines anderen Genotyps. Das bedeutet, dass Genotypen mit mehr Nachkommen eine größere Fitness haben. Einen guten Überblick zu „Fitness“ und „Genotypen“ findet man in Wikipediaw1.
Genom: Die gesamte DNA eines Lebewesens. Darunter versteht man üblicherweise die DNA im Zellkern, im Gegensatz zur DNA in Mitochondrien, oder in Plastiden. Nähere Informationen im Artikel ‘What is a genome’ auf der Homepage des US National Library of Medicinew2.
Positive Selektion: Die natürliche Selektion ist einer der Evolutionsmechanismen. Sie beschreibt die unterschiedlichen Überlebens- und Vermehrungsraten von Lebewesen unter bestimmten Umweltbedingungen. Sie ist „positiv“, wenn bestimmte Merkmale das Überleben und die Reproduktion ihrer Träger verbessern.
Selektionsdruck: Umweltbedingungen (z.B. Temperatur, Anwesenheit von Parasiten, räuberisches Verhalten oder Aggression von Mitgliedern derselben Art), die unterschiedliche Überlebens- und Vermehrungschancen einzelner Individuen verursachen.
Merkmal: eine oder mehrere Eigenschaften eines Lebewesens (z.B. Körpergröße, Resistenz gegenüber Antibiotika, die Fähigkeiten, Farben zu sehen oder die Zunge zu rollen).
Acknowledgements
The author is grateful to David Hughes, Mehmet Somel and Ania Lorenc for helpful comments on the article.
References
- Harris EE, Meyer D (2006) The molecular signature of selection underlying human adaptations. American Journal of Physical Anthropology 131(S43): 89-130. doi: 10.1002/ajpa.20518
- Der Artikel gibt einen guten Überblick über den Forschungsstand der molekularen Evolution im Menschen.
- Hoekstra H et al. (2006) A single amino acid mutation contributes to adaptive beach mouse color pattern. Science 313: 101-104. doi: 10.1126/science.1126121
- Dieser und andere Artikel zur Fellfarbe von Mäusen der Hopi Hoekstra’s Forschungsgruppe sind auf der Homepage der Harvard Universität abrufbar: www.oeb.harvard.edu/faculty/hoekstra/Links/PublicationsPage.html
- Weitere Informationen im Folgeartikel, in dem die Entdeckung von Agouti, einem negativen Regulator des Beitrags von McR1 zur Anpassung der Fellfarbe in Peromyscus beschrieben wird:
- Steiner CC, Weber JN, Hoekstra HE (2007) Adaptive variation in beach mice produced by two interacting pigmentation genes. PLoS Biology 5: e219. doi: 0.1371/journal.pbio.0050219
- Dieser und andere Artikel in PLoS Biology können kostenlos heruntergeladen werden.
- Der folgende Artikel fasst die adaptive Färbung von Wirbeltieren zusammen:
- Hoekstra HE (2006) Genetics, development and evolution of adaptive pigmentation in vertebrates. Heredity 97: 222-234. doi: 10.1038/sj.hdy.6800861
- Der Artikel kann kostenlos von der Homepage des Heredity Journals heruntergeladen werden: www.nature.com/hdy
- Ein Überblick über Hopi Hoekstra’s neueste Forschungsergebnisse gibt es im Blog von John Hawks: http://johnhawks.net/weblog/topics/evolution/selection/hoekstra-2009-adaptive-pigmentation.html
- Lamason RL et al. (2005) SLC24A5, a putative cation exchanger, affects pigmentation in zebrafish and humans. Science 310: 1782-1786. doi: 10.1126/science.1116238
- Mwangi MM et al. (2007) Tracking the in vivo evolution of multidrug resistance in Staphylococcus aureus by whole-genome sequencing. Proceedings of the National Academy of Sciences of the United States of America 104: 9451-9456. doi: 10.1073/pnas.0609839104
- Perry GH et al. (2007) Diet and the evolution of human amylase gene copy number variation. Nature Genetics 39: 1256-1260. doi: 10.1038/ng2123
- Eine Überblick über Panda’s Forschung findet man auf: http://pandasthumb.org/archives/2008/12/amylase-and-hum.html
- Stefansson H et al. (2005) A common inversion under selection in Europeans. Nature Genetics 37: 129-137. doi: 10.1038/ng1508
- Eine Zusammenfassung der Arbeit findet sich auch bei Evolgen: http://evolgen.blogspot.com/2005/02/human-inversion-under-selection.html
- Tishkoff SA et al. (2006) Convergent adaptation of human lactase persistence in Africa and Europe. Nature Genetics 39: 31-40. doi: 10.1038/ng1946
- Eine Zusammenfassung der Forschungsergebnisse findet sich auch in der New York Times: www.nytimes.com/2006/12/10/science/10cnd-evolve.html?_r=1
Web References
- w1 – Ein gutter Überblick über die Begriffe ‘Fitness’ and ‘Genotype’, bietet Wikipedia: http://en.wikipedia.org/wiki/Fitness_(biology) und http://en.wikipedia.org/wiki/Genotype
-
w2 – Weitere Informationen zu Genomen und dem Humangenomprojekt, siehe ‘What is a genome’ auf der Webseite des US National Library of Medicine: http://ghr.nlm.nih.gov/handbook/hgp/genome
Resources
- Falls Sie dieser Artikel interessiert hat, könnten auch andere Artikel aus Science in School von Interesse sein:
- Haubold B (2010) Review of Why Evolution is True. Science in School 14. www.scienceinschool.org/2010/issue14/evotrue
- Leigh V (2008). Interview mit Steve Jones: die Bedrohung durch den Kreationismus. Science in School 9. www.scienceinschool.org/2008/issue9/stevejones/german
- Patterson L (2010) Getting ahead in evolution. Science in School 14: 16-20. www.scienceinschool.org/2010/issue14/amphioxus
- Pongsophon P, Roadrangka V and Campbell A (2007) Counting Buttons: demonstrating the Hardy-Weinberg principle. Science in School 6: 30-35. www.scienceinschool.org/2007/issue6/hardyweinberg
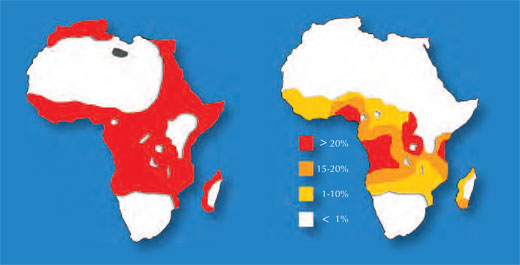
- Weitere Informationen zur Malaria:
- Hodge R (2006) Fighting malaria on a new front. Science in School 1: 72-75. www.scienceinschool.org/2006/issue1/malaria
- Weitere Informationen zur Struktur von Stärke, die durch die Speichelamylase abgebaut wird:
- Cornuéjols D (2010) Starch: a structural mystery. Science in School 14: 22-27. www.scienceinschool.org/2010/issue14/starch
Review
Der Artikel beschreibt eine Reihe interessanter Beispiele adaptiver Evolutionsprozesse auf molekularer Ebene. Dabei werden die Schwierigkeiten bei der Aufklärung des Zusammenhangs zwischen adaptiven DNA-Sequenzen und der individuellen Fitness beim Menschen und die Notwendigkeit von Medellorganismen aufgezeigt.
Der Artikel bietet eine exzellente Grundlage zur Bearbeitung übergeordneter Fragestellungen, die das Verständnis der natürlichen Selektion und der Fitness im Menschen und in Modellorganismen ermöglichen. Einige Beispiele:
- Erkläre die Prozesse, die für die natürliche Selektion verantwortlich sind..
- Was vesteht man unter dem Begriff „Fitness“?
- Erkläre, wie das Sichelzellanämieallel einen selektiven Vorteil für manchen menschlichen Populationen bewirkt.
- Welche Probleme treten auf, wenn man kausale Zusammenhänge zwischen adaptiven DNA-Sequenzen und der Fitness herstellen will?
- Konstruiere ein Flussdiagramm, das den adaptiven Wert der Fellfarbe der Alabama-Küstenmaus erklärt.
- Wie konnten die Wissenschafter/innen einen Zusammenhang zwischen genetischen Veränderungen in Staphylococcus aureus und der Antibiotikaresistenz herstellen?
Der Artikel erlaubt es Schüler/innen sich mit dem Zusammenhang zwischen DNA, Aminosäuresequenz, Proteinstruktur und – funktion bei der Sichelzellanämie zu beschäftigen. Der Text eignet sich, um Klassendiskussionen über die Methoden und Probleme bei der Untersuchung der molekularen Basis evolutionärer Beziehungen und den ethischen Fragen bei der Untersuchung menschlicher Populationen zu stimulieren. Es könnten fachübergreifende Arbeiten zur Wissenschaftsgeschichte und populationsgenetischer Evolutionsstudien durchgeführt werden.
Mary Brenan, Großbritannien